Часть 1 Современные узконосые обезьяны Демонстрационная версия (до консультаций со специалистами, корректировки карт, проведения тщательного поиска лучших фотографий и т.д.)
Информация для данной презентации бралась из русской и английской Wikipedia и с сайта Primat conservancy.
Зеленым шрифтом отмечены оригинальные тексты автора не прошедшие научного редактирования.
Фиолетовым жирным шрифтом отмечены вопросы к специалистам-приматологам. Эти вопросы часто сформулированы в виде утверждений, но фиолетовый цвет предполагает консультацию со специалистами.
Данная презентация является, насколько нам известно (очень вероятно, что это не так, и это надо проверить), первой попыткой представить географию расселения всех узконосых обезьян в одном проекте в одном дизайне и провести первичный анализ полученной картины.
Панели общей длиной 130 м выставлены в Парке Леко.
Классификация современных узконосых обезьян
Упрощенная систематика. Нам кажется, что для БТК должны быть четко определены категории "суперсемейство", "надсемейство" и "семейство". Должна быть определена входимость каждого рода во все категории без пропусков в иерархии. Например, род Pongo должен входить в семейство Pongo, несмотря на то, что он в этом семействе единственный. В случае единственного рода в семействе, имя семейства такое же, как имя рода. Если вообразить, что в будущем род Pongo будет разделен на 2 рода, то новый род будет входить в уже имеющееся семейство (ну и в него должны входить вымершие рода).
Наш вид Хомо сапиенс является единственным современным видом в роде Хомо, который входит в семейство Хомо, которое входит в надсемейство Человекообразных, которое входит в суперсемейство Узконосых обезьян.
Всего узконосых обезьян 29 родов, включающие 183 вида (не учитывая Хомо сапиенс).
Понятие рода и вида, принцип Гауза
Wikipedia: Род -- вторая главная категория в биологической систематике, следует за категорией вид.
В отличие от вида, где с оговорками и исключениями к одному виду относятся особи способные скрещиваться и давать плодовитое потомство, для рода нет никакого общего критерия. Однако, анализ существующей, в основном сложившейся еще до молекулярной генетики, схемы объединения видов узконосых обезьян в рода позволяет дать следующее базовое содержательное определение: Род -- совокупность близкородственных видов занимающих одну экологическую нишу. Это упрощенный (не учитывающих некоторые случаи родообразования, о которых ниже) и несколько неожиданный вариант определения рода, т.к. исторически систематика происходила в основном по внешним признакам и по образу жизни, позже по генетическим данным, без специального анализа экологических ниш. Проведенный нами анализ корреляции современной систематики с географией расселения и с распределением по экологическим нишам, позволяет увязать дефиниции "род" и "экологическая ниша" в одном определении.
Экологические ниши определяются следующим (полным или неполным) набором параметров: -- климатические условия в достаточно широких пределах, -- ландшафт -- саванна, джунгли, высокогорье и т.д., -- высотный уровень -- поверхность земли, нижний ярус, средний ярус, верхний ярус деревьев, -- рацион питания -- насекомые, фрукты, орехи, соки растений, листья и т.д., -- взаимодействие с хищниками.
Принцип Гауза: Два вида, которые конкурируют за один и тот же ограниченный ресурс, не могут стабильно сосуществовать. Когда один вид имеет хотя бы малейшее преимущество перед другим, тот, у кого это преимущество, будет доминировать в долгосрочной перспективе. Это приводит либо к вымиранию более слабого конкурента, либо к сдвигу в сторону другой экологической ниши.
Из принципа Гауза следует, что ареалы видов одного рода зависимы друг от друга, они не пересекаются (но могут иметь общие границы), а ареалы родов могут практически независимо накладываются друг на друга. Этому следствию из принципа Гауза соответствует наблюдаемое на картах расселение узконосых обезьян и наше "экологическое" определение рода.
Можно сформулировать упрощенный (приемлемый для БТК) критерий видов одного рода для мысленного эксперимента: если любой вид переселить в ареал другого вида (предварительно выселив этот другой вид), то он сможет адаптироваться к новым условиям.
В презентации мы приводим карты расселения родов и видов узконосых обезьян, даем краткие упрощенные формулировки экологических ниш для всех родов, анализируем причины случаев несоответствия карт принципу Гауза, т.е. случаи когда ареалы видов одного рода пересекаются. Если карты не противоречат принципу Гауза, то называем эту ситуацию соответствующей принципу Гауза.
При анализе родов узконосых обезьян, принцип Гауза явно нарушается только в двух случаях: 1. Для рода Cercopithecus Мартышки. Но подробный анализ этого рода, в котором имеют важнейшее значение поливидовые ассоциации, позволяет сделать предположение, что виды входящие в поливидовые ассоциации могли бы быть объединены в группы более высокого ранга, чем виды. Вероятно, было бы правильным объединить некоторые виды этого рода в самостоятельные рода (на панеле Мартышки это рассмотрено подробно и сформулированы вопросы к специалистам-приматологам). 2. Для рода Macaca Макаки. Скорее всего информации полученной нами из Wikipedia и сайта Primat conservancy не достаточно для определения экологических ниш макак (на панеле Макаки сформулированы вопросы к специалистам-приматологам).
Структура панелей
Мы следуем структуре сайта Primat conservancy. Одна страница у них -- одна панель у нас. В каждой панеле объединены рода одного семейства (или таксона высшего по отношению к родам).
Для семейств мы даем общую карту родов с описанием распределения родов по экологическим нишам.
Далее идут карты видов по каждому роду. Слева от каждой карты по роду показаны группировки видов по визуальной похожести (пока очень приблизительно, нужно сделать более тщательно). Рядом с картой рода дается формулировка экологической ниши рода, анализируются границы видов. Далее следуют общие характеристики для всех видов рода (это сделано пока не достаточно аккуратно, будут уточнения после более качественной обработки имеющихся данных и консультаций со специалистами).
Для подробного анализа ландшафтов нами делаются (пока сделаны только на картах толстотелов) подробные границы на Google-картах, там можно приближать и видеть ландшафт в режиме "спутник" с очень высоким разрешением -- вплоть до отдельных деревьев.
Далее дополнительные материалы. У каждого вида собственный раздел, в котором приведены интересные материалы, лучшие и самые качественные найденные фотографии, видео-ролики по QR-кодам, политические карты со сравнением численности с численностью Хомо сапиенс, список зоопарков и т.п. Эти материалы будут со временем (постоянно) совершенствоваться и дополняться.
Филогенетические деревья современных родов и видов обезьян
ФГД пока не показаны (в Интернете нет этого контента). После консультаций со специалистами ФГД предполагается показать для каждого рода. Это в некоторых случаях приведет к изменениям в формулировках экологических ниш и в объяснениях особенностей расселения.
Здесь квинтэссенция этой статьи в упрощенных формулировках автора презентации с дополнениями, чтобы охватить все возможные способы видообразования:
1. Аллопатрическое -- географическая изоляция. 2. Перипатрическое -- частный случай аллопатрического, когда некоторые группы оказываются изолированными в приграничье большого ареала (важное значение может иметь дрейф генов).
3. Парапатрическое -- большой ареал имеет некоторый градиент экологических отличий, который приводит к направленным модификациям вида, процесс видообразования завершается под действием фактора избирательности скрещивания.
4. Симпатрическое -- особи вида в одном ареале находят более специализированные экологические ниши, адаптируются к ним, а особи не приспособившиеся к специализированным нишам имеют меньшую приспособленность и постепенно вид разделяется на два.
5. Гибридизация -- виды с одним кариотипом (числом, размером, формой и т.д. хромосом), даже при существенном различии генотипов, иногда могут дать плодовитое потомство.
6. "Кариотипическое" -- одинаковое изменение кариотипа одновременно у мужской и женской особи исключающее возможность давать плодовитое потомство с остальными особями вида, но позволяющее дать плодовитое потомство при скрещивании между собой.
7. "Ароморфическое" -- мутация у одной особи дающая существенное преимущество и поддерживаемая половым отбором (пример, на фрагменте филогенетического дерева приматов (гиперссылка) -- появление бесхвостых обезьян, ставших позже человекообразными).
Родообразование
Рода образуются в результате 3-х категорий событий:
1. Аллопатрическое или перипатрическое видообразование состоявшееся достаточно давно, чтобы произошло разделение не только на два вида, но и изменения дошли до столь явных отличий, что новые виды очень существенно отличаются по ряду признаков. 2. Симпатрическое видообразование состоявшееся достаточно давно, чтобы произошло устойчивое разделение не только на два вида. Но с позиций нашего "экологического" определения рода ситуация должна быть описана по другому. Момент состоявшегося симпатрического видообразования нужно считать моментом разделения рода на два. Т.е. образование двух видов и двух родов при симпатрическом видообразовании -- одно и тоже событие. Ареалы таких родов обычно пересекаются. 3. "Мгновенное" видообразование ("кариотипическое" и "араморфическое") -- новый вид (уже сразу род) выживет, если покинет ареал старого вида или вытеснит старый вид или им удастся разделить ареал.
Гипотеза KF-видообразования
На картах расселения узконосых обезьян, наиболее интересно, кроме почти всегда очевидных случаев аллопатрического и перипатрического видообразования, рассмотреть оставшиеся случаи. Трудно не сделать вывод, что картина расселения видов многих родов узконосых (сейчас мы начали делать аналогичную работу по широконосым, посмотрим, как будет там) часто может быть объяснена только парапатрическим видообразованием.
Причем, это парапатрическое видообразование, кажется, имеет какой-то мощный фактор усиления. Классическим "спокойным" вариантом парапатрического видообразования трудно объяснить, например, карту расселение макак на Сулавеси, больше похожую на политическую карту.
Предложим следующую гипотезу, которую назовем гипотезой KF-видообразования. Кратко смысл такой. Дарвиновская эволюция, почему-то, поддерживает "ген ксенофобии". В результате особи удаленных популяций вида, которые вполне хорошо могут скрещиваться, но уже имеют насколько-то различный фенотип (начальная стадия парапатрического видообразования) начинают скрещиваться меньше, чем могли бы. Предполагаем, что из-за ксенофобии. В результате происходит дробление видов.
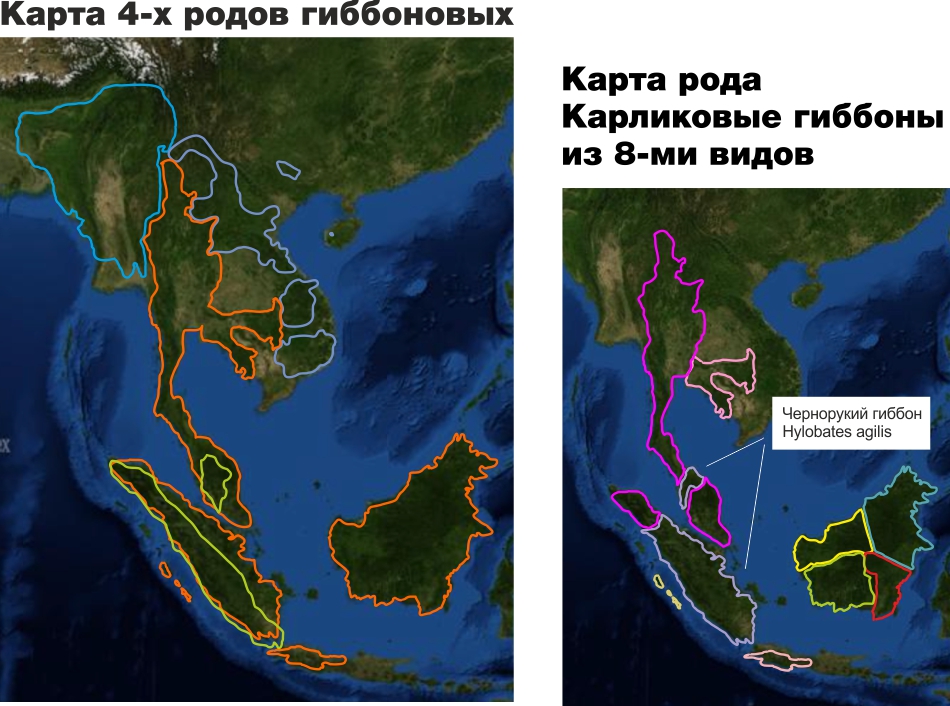
В зоопарках виды всегда представляются примерно так (в дополнение к нижеизложенной критике отметим, что не аккуратный дизайн верхней части в этом конкретном случае не позволяет понять, где официальное наименование вида, включающее официальное название рода, а где неофициальное название вида на английском языке)
Фактически это дезинформация. Неспециалист (не проведя специального самостоятельного исследования в интернете) не поймет, что гиббоны проживают на гораздо более обширной территории, и чернорукий гиббон является одним из 8-ми видов рода карликовых гиббонов, одного из 4-х родов гиббоновых и представляет в Московском зоопарке все семейство гиббоновых.
Для экскурсоводов, студентов биофаков, специалистов и всех заинтересованных в изучении зоологии могли бы использоваться такие планшеты
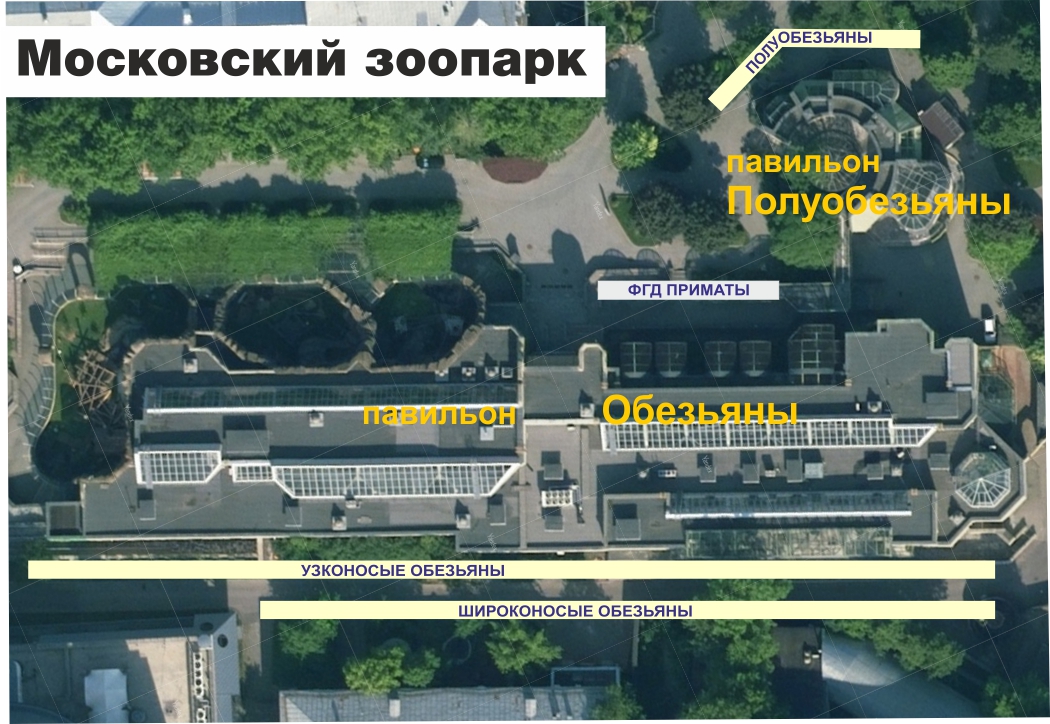
В зоопарках можно было бы выставлять информацию оформленную аналогично нашей презентации. Например, в Московском зоопарке -- вдоль задней аллеи павильона обезьян панели узконосых и широконосых, а полуобезьян рядом с их павильоном.
В идеале такие панели должны быть для всех животных, хотя бы в электронном формате на планшете.
Автор -- Константин Могильницкий Дизайн -- Ольга Кутукова