|
Предложим следующую гипотезу, которую назовем гипотезой KF-видообразования. Кратко смысл такой. Дарвиновская эволюция, почему-то, поддерживает "ген ксенофобии". В результате, особи удаленных популяций вида, которые вполне хорошо могут скрещиваться, но уже имеют насколько-то различный фенотип (начальная стадия парапатрического видообразования) начинают скрещиваться меньше, чем могли бы. Предполагаем, что из-за ксенофобии. В результате происходит дробление видов.
Ниже приведены аргументы в пользу KF-видообразования.
Мимикрия под аллопатрическое видообразование
Река разделила популяцию. Прошло 3 тыс. лет. Река уже уменьшилась, возможно смешение популяций, но механизм KF-видообразования продолжает работать на создание разных видов. Внешне это может выглядеть, как аллопатрические видообразование, т.к. есть четкая граница по реке.
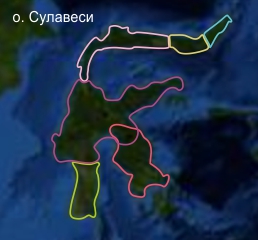
Пример с картами для колобусов. На картах родов мы размещает QR-коды на гугловские карты, где нами намечены и уточняются границы ареалов. Сплошная линия -- граница взятая из из русской и английской Wikipedia или с сайта Primat conservancy. Пунктирная -- уточненная нами по наблюдаемому при сильном приближении ландшафту (будут еще уточнения после консультаций со специалистами). Пока сделано только для колобусов.
Рассмотрим несколько фрагментов границы разделения видов (1) и (2)

Река широкая, но есть места, где острова и деревья близко, трудно представить, что тысячи лет колобусы не могут ее преодолеть.

Мимикрия под симпатрическое видообразование
Река разделила популяцию. Прошло 3 тыс. лет. Река уже уменьшилась или почти исчезла. Смешению популяций ничего не мешает, но механизм KF-видообразования продолжает работать на создание разных видов. Постепенно виды начинают питаться немного разной пищей (можно допустить, что есть неоднородность в распределение растений с двух сторон реки). Внешне это может выглядеть, как симпатрическое видообразование.
Аналогия с половым отбором
Особи имеют специальные предпочтения при выборе полового партнера.
Оптимальные отличия в генотипе скрещивающихся самцов и самок
Какое оптимальное различие генотипов (приматов или в целом млекопитающих) для лучшей приспособленности?
Групповое поведение препятствует близкородственному скрещиванию.
Какой механизм препятствования гибридизации подвидов?
Роль альфа-самцов и будущих альфа-самцов
Стремящийся захватить группу потенциальный новый альфа-самец встречает большее подозренее, если имеет дополнительные фенотипические отличия.
Защита территории вида усиливает парапатрию
Особи имеющие ген ксенофобии более территориальны. Защита территории происходит с более отчаяно.
Защищающиеся группы лучше знают территорию на которой происходит конфликт, что при прочих равных делает вероятность отстоять территорию выше, чем захватить.
Почти все узконосые территориальны, и в случае появления более отличных фенотипически групп, могут вести себя предельно агрессивно.
Почему дарвиновская эволюция "любит" KF-видообразование
1. Мартышки-гусары при резком изменение условий вымрут с большей вероятностью, чем павианы или зеленые мартышки, т.к. суммарный генофонд мартышек-гусар меньшего объема.
2. При прочих равных красные колобусы имеющие большее количество видов, выживут с большей вероятностью, чем черно-белые или оливковый.
3. Повышение скорости видообразования способствует расширению ареала. Сравним ареал мартышек-гусаров и павианов и зеленых мартышек.
Граничащие подвиды
Мы не делали анализ по подвидам. Аллапатрически или перипатрически раздробленные виды, мы просто рассматриваем, как различные популяции. В отличие от этого, граничащие подвиды с точки зрения парапатрического видообразования очень интересны. Вероятно, мы проведем такой анализ ландшафтов их ареалов позже.
Разделится ли Хомо сапиенс на разные виды
На этот вопрос некоторые ученые отвечают отрицательно. Происходит же обмен генами между всеми популяциями людей.
Попробуем написать формулу этого процесса для 2-х популяций имеющих стабильную численность.
Очевидно работают следующие параметры:
-- N1 и N2 -- численность популяций,
-- g -- скорость мутагенеза (кол-во мутаций на одно поколение), оно одинаково для обеих популяций,
-- k -- количество межпопуляционных скрещиваний на одно поколение дающих фертильное потомство,
Для сапиенсов, имеющих культурные различия в разных популяциях, эти различия имеют дополнительное значение для активации гена ксенофобии.
Скорее всего параметры N1 и N2 сократятся и ответ на вопрос произойдет ли видообразование будет зависеть от преобладания параметра g над k.
Понятно, что когда популяций много и скрещивание возможно между всеми, то формула резко усложнится, но параметры g и k в ней останутся и продолжат действовать в тех же направлениях.
Далее, мы не расписываем, такая формула, наверняка уже существует. Интересно ее применение к популяциям приматов и, в частности, к сапиенсам.
|