Целями презентации являются получение общей картины географического, экологического и фенотипического разнообразия приматов, изучение на отдельном отряде млекопитающих природы видообразования и родообразования.
Информация о современных приматах -- из Wikipedia и с Primat conservancy.
Оригинальные термины, тексты автора не прошедшие научного редактирования и вопросы к специалистам-приматологам отмечены курсивом. Эти вопросы часто сформулированы в виде утверждений, но курсив предполагает необходимость консультации со специалистами.
Презентация является, насколько нам известно (вероятно, что это не так, и это надо проверить), первой попыткой представить географию расселения всех приматов в одном проекте в одном дизайне и провести первичный анализ полученной картины.
Панели общей длиной 280 м выставлены в Парке Леко.
Экологическая ниша -- место, занимаемое видом в природной системе. Экологические ниши определяются следующим (полным или неполным) набором параметров:
ландшафт -- саванна, джунгли, высокогорье и т.д., питание -- всеядность, насекомоядность, фрукты, орехи, соки растений, листья и т.д., высотный уровень -- поверхность земли, нижний, средний, верхний ярус леса,
взаимодействие с хищниками, климатические условия в достаточно широких пределах.
Приматы -- отряд млекопитающих отделившийся от отряда грызунов, в соответствии со специализацией -- древесные тропических лесов. Параллельная ветвь -- грызуны-беличьи -- древесные смешанных и хвойных лесов. С точки зрения экологической ниши, можно определить приматов, как древесных млекопитающих населяющих лиственные вечнозеленые леса или имеющих предков с такой экологической нишей.
Род (Wikipedia) -- вторая главная категория в биологической систематике, следует за категорией вид. В отличие от вида, где с оговорками и исключениями к одному виду относятся особи способные скрещиваться и давать плодовитое потомство, для рода нет никакого общего критерия. Однако, анализ существующей, в основном сложившейся еще до молекулярной генетики, схемы объединения видов приматов в рода позволяет дать следующее базовое содержательное определение: Род -- совокупность близкородственных видов занимающих одну экологическую нишу. Это упрощенный (не учитывающий некоторые случаи родообразования, о которых ниже) и несколько неожиданный вариант определения рода, т.к. исторически систематика происходила в основном по внешним признакам и по образу жизни, позже по генетическим данным, но без специального анализа экологических ниш родов с учетом географии их ареалов. Проведенный нами анализ корреляции современной систематики с географией ареалов и с распределением по экологическим нишам, позволяет увязать дефиниции "род" и "экологическая ниша" в одном определении.
Принцип Гауза: Два вида, которые конкурируют за один и тот же ограниченный ресурс, не могут стабильно сосуществовать. Когда один вид имеет хотя бы малейшее преимущество перед другим, тот, у кого это преимущество, будет доминировать в долгосрочной перспективе. Это приводит либо к вымиранию более слабого конкурента, либо к сдвигу в сторону другой экологической ниши. Принцип Гауза можно переформулировать: ареалы видов одного рода зависимы друг от друга, они не пересекаются (но могут иметь общие границы), а ареалы родов могут практически независимо накладываются друг на друга. Принципу Гауза соответствует наблюдаемое на картах расселение приматов и наше "экологическое" определение рода. Можно сформулировать упрощенный (приемлемый для БТК) критерий видов одного рода: если в мысленном эксперименте любой вид переселить в ареал другого вида того же рода (предварительно выселив этот другой вид), то он сможет адаптироваться к новым условиям и экосистема при этом не претерпит принципиальных изменений.
Следствие из принципа Гауза (понятие введенное нами, вероятно, есть какой-то общепринятый аналог): Род всегда займет весь географически связанный с его местонахождением ареал соответствующий его экологической нише. Это следствие опирается на принципиально экспоненциальный рост численности каждого вида при отсутствие ограничений. Если покрыт не весь экологически возможный и при этом географически доступный ареал, то надо искать причину, почему возникло такое ограничение.
Соответствие принципу Гауза и следствию из принципа Гауза. В презентации для каждого из 4-х связанных глобальных ареалов приматов (Африка, Азия, Южная Америка, Мадагаскар) показаны карты расселения родов и расселение родов проанализировано на соответствие принципу Гауза и следствию из принципа Гауза. Затем идут подробные разделы по семействам с картографией по родам и с анализом соответствия расселения видов принципу Гауза.
Способы видообразования
Статья о видообразовании в Wikipedia. Здесь квинтэссенция этой статьи в упрощенных формулировках автора презентации с дополнениями, чтобы охватить все возможные способы видообразования:
Аллопатрическое -- географическая изоляция. Перипатрическое -- частный случай аллопатрического, стратовой ситуацией является изоляция группы в приграничье первоначально общего ареала (важное значение может иметь дрейф генов). Парапатрическое -- большой ареал имеет некоторые градиенты экологических отличий, который приводит к направленным модификациям вида или существует какой-то ландшафтный рубеж, который преодолим, но с затруднениями или что-то подобное, в результате чего возникает нежесткое разделение на популяции, далее возникают небольшие фенотипические отличия, процесс видообразования завершается под действием фактора избирательности скрещивания. Симпатрическое -- особи имеющие какое-то выраженное фенотипическое отличие (изменчивость) находят соответствующие более специализированные экологические ниши, адаптируются к ним, и постепенно вид разделяется на два. Такой же результат, если естественный отбор не благоприятствует среднему проявлению фенотипа относительно фенотипов полярных изменчивостей. Гибридизация -- виды с одним кариотипом (числом, размером, формой и т.д. хромосом) даже при существенном различии генотипов, иногда могут дать плодовитое потомство. Кариотипическое -- одинаковое изменение кариотипа одновременно у мужской и женской особи исключающее возможность давать плодовитое потомство с остальными особями вида, но позволяющее дать плодовитое потомство при скрещивании между собой. Ароморфическое -- мутация у одной особи дающая существенное преимущество и поддерживаемая естественным или (и) половым отбором (пример -- на филогенетическом дереве приматов -- появление бесхвостых обезьян, ставших позже человекообразными).
Родообразование
Виды, имея одинаковую экологическую нишу, могут долгое или короткое время существовать в пределах одного рода. Внутри этого рода виды могут расширять за счет других видов свои ареалы, какие-то виды могут вымирать, могут появляться новые. В результате 3-х категорий событий могут появляться новые рода:
Аллопатрическое, перипатрическое или парапатрическое видообразование состоявшееся достаточно давно, чтобы произошло разделение не только на два вида, но и изменения дошли до столь явных отличий, что виды очень существенно отличаются по ряду признаков. Симпатрическое видообразование состоявшееся достаточно давно. Но с позиций "экологического" определения рода ситуация должна быть описана по другому. Момент состоявшегося симпатрического видообразования нужно считать моментом разделения рода на два. Т.е. образование двух видов и двух родов при симпатрическом видообразовании -- одно и тоже событие. Мгновенное видообразование ("кариотипическое" и "араморфическое") -- новый вид (уже сразу род) выживет, если покинет ареал старого вида или вытеснит старый вид.
Графика на филогенетическом дереве
Событиями родообразования мы называем события, которые приводят к появлению новых родов, меняют экологические ниши родов, приводят к исчезновению родов.
Представление событий родообразования на ФГД (филогенетическом дереве)
Симпатрическое ответвление -- ответвление с наклоном Например, событие ответвления ведущего к шерстокрыловым. Способность при прыжках вниз преодолевать максимальное горизонтальное расстояние способом планирования могло развиваться одновременно у многих видов тупаевых. Начинается процесс отделения от прочих тупаевых. Постепенно побеждает одна ветвь. Идет видообразование, вымирания видов, совершенствование. На ФГД это показывается только одной веткой с наклоном символизирующем совокупность вышеописанных процессов.
Стабилизирующая замена -- прямая со сменяющими друг друга без ответвлений родами Предполагается, что причиной вымирания родительского рода являются потомки более приспособленные в той же экологической нише. Экологическая ниша при этом может относительно слабо меняться расширяясь, сужаясь, отклоняясь в зависимости от внешних факторов и от достигнутого потомками совершенствования признаков.
Аллопатрическое ответвление (миграция) -- вертикальное ответвление В каждом случае необходимо описание способа миграции и карта миграции.
Аллопатрическое разветвление -- вертикальные вверх и вниз ответвления В каждом случае необходимо описание конкретного географического события и карта разделения ареала. Например, событие разделения шерстокрыловых на рода. 22 Ма Индонезия разделилась на острова. Произошло аллопатрическое видообразование. К нашему времени шерстокрылы из аллопатрически возникших видов морфологически и генетически изменились настолько, что признаны разными родами, хотя экологические ниши у этих родов одинаковые.
Парапатрическое родообразование -- расходящиеся наклонные от карты предкового рода к картам новых родов
Без специальных объяснений -- все видно по картам.
Ароморфическое ответвление -- вертикальное ответвление Описание мутационного события и причин его поддержки естественным или (и) половым отбором, приводящего к мгновенному ответвлению нового рода.
Вымирание -- крестик Причины вымирания прописываются ближе к крестику.
Современное незаконченное событие родообразования -- в рамке показыается не род, а вид, используются зеленые линии. Подпись: "Текущий процесс родообразования".
Биологические смыслы
Каждое событие родообразования должно быть сопровождено формулировкой биологического смысла. Если биологический смысл доподлинно не известен, то должен быть указан предполагаемый (самый вероятный или самый ясный). На уровне БТК альтернативы рассматриваться не должны, кроме отдельных случаев, где само существование альтернатив может быть интересным знанием уже на базовом уровне.
Пиктограммы карт на ФГД
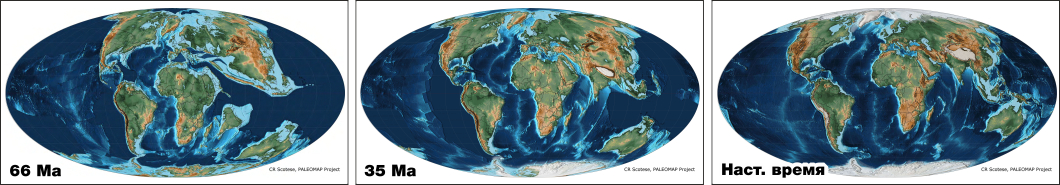
Палеогеография сильно отличается от современной географии. Однако контуры континентов и их взаимное расположение в период эволюции млекопитающих (последние 66 млн. лет представленные на ФГД) уже достаточно близки к современным.
На пиктограммах карт, соответствующих времени появления родов приматов, использованы современные контуры суши. Условно считается, что джунгли и саванны находились там же, где сейчас. Современный ареал заливается красным цветом. Если ареал рода был принципиально большим, чем в наше время, то потерянный ареал заливается темно-синим цветом, текстом указывается причина сокращения ареала.
Термины и условные обозначения в таблицах
Соотношение веса самки к весу самца -- еще можно назвать коэффициентом полового весового диморфизма. Например, для нашего вида Хомо сапиенс средний вес мужчин в мире (по данным на 2020 год) -- 80 кг, женщин -- 66 кг. Таким образом, коэффициент весового полового диморфизма у нас около 0,83.
Хвост, % от длины тела -- если 100%, то длина хвоста равна длине остального тела.
Площадь группы, км2 -- площадь, которая занимает группа. Часто самки и дети занимают не всю площадь. У нас дано значение для площади, которую контролируют самцы.
Натальная группа -- это группа, в которой животные находятся с момента рождения. Если по достижении половозрелости группу покидают самцы -- "м" , если самки -- "ж". Если самки остаются в своих натальных группах, то они состоят в матрилинейном родстве, в этом случае самцы уходят, чтобы присоединиться к новым группам или сформировать их по достижении совершеннолетия.
Описание последовательности и метода исследования
1. Основываясь на данных русской и английской Wikipedia и сайта Primat conservancy по ареалам видов создавалась единая карта каждого рода. Анализировалось соответствие расселения видов принципу Гауза. Анализ экологических ниш проводился по траектории: ландшафт -- ярус леса -- сезонное питание с учетом размерного класса -- несезонное питание -- разделение дневных и ночных. Если после этого ниши пересекаются, то проводится анализ по хищникам и другим аспектам. Если (в редких случаях) не удавалось однозначно определить экологическую нишу особенную для конкретного рода, то проблема фиксировалась в виде вопроса курсивом. Дополнительно карты по родам каждого семейства сводились в одну карту, проводилось дополнительное уточнение соответствия принципу Гауза и следствию из принципа Гауза, уточнялись экологические ниши, выявлялись случаи аллопатрического и парапатрического разделения родов.
2. Для каждой части света (для приматов части света это Африка, Азия, Южная Америка и Мадагаскар) карты по родам сводились в группы с близкими экологическими нишами независимо от филогенетической близости (на одну карту могли даже попасть обезьяны и полуобезьяны, если у них близкие экологические ниши). Дополнительно анализировалось соответствие принципу Гауза и следствию из принципа Гауза. Окончательно уточнялись экологические ниши. 3. На основании книги С. Дробышевского "Кайнозой" было создано ФГД приматов в первом приближении. 4. На основании ФГД в первом приближении и разработок по определению экологической ниши каждого рода строилось логически непротиворечивое ФГД с определением места на нем каждого рода. На ФГД показаны все современные рода приматов.
Интересные ожидаемые и неожиданные результаты
Удивительно, что столь простой, использованный нами, метод исследования является в основном достаточным для определения экологических ниш, что свидетельствует об ограниченном их количестве и о полноте их занятия. Нет экологически возможных и не занятых ниш. Одни и те же ниши в разных частях света заняты самыми разными и филогенетически очень далекими родами приматов.
Экологических ниш довольно небольшое и конкретное количество. Ниши более обширны, чем можно было предполагать, например, не нашло подтверждения предположение, что разные рода листоядных в одном ареале питаются листьями разных деревьев, такого относительно узкого разделения ниш нет. Практически нет случаев, где бы экологическая ниша определялась хищниками (исключением, вероятно, являются черно-белые и красные колобусы). Экологические ниши определяются конкуренцией за пищу, т.е. являются почти строго пищевыми.
Почти не обнаружено случаев несоответствия принципу Гауза при анализе видов одного рода. Например, при анализе родов узконосых обезьян, принципу Гауза не соответствуют только два случая: 1. Для рода Cercopithecus Мартышки. Но подробный анализ этого рода, в котором имеют важнейшее значение поливидовые ассоциации, позволяет сделать предположение, что виды входящие в поливидовые ассоциации могли бы быть объединены в группы более высокого ранга, чем виды. Вероятно, было бы правильным объединить некоторые виды этого рода в самостоятельные рода (на панеле Мартышки это рассмотрено подробно и сформулированы вопросы к специалистам-приматологам). 2. Для рода Macaca Макаки. Скорее всего информации полученной нами из Wikipedia и сайта Primat conservancy не достаточно для определения экологических ниш макак (на панеле Макаки сформулированы вопросы к специалистам-приматологам).
Следствие из принципа Гауза нарушается только для родов: мандрилы, гелады, мартышки-гусары, отолемуры. В каждом случае приведено объяснение (если нам удалось найти объяснение в источниках) или высказано предположение об этом несоответствии (если мы не нашли объяснения в источниках).
Антропогенное воздействие на приматов на уровне видов очень существенно, однако при взгляде на уровне родов нарушение следствия из принципа Гауза незначительно. Рода заселяют практически все соотвествующие их экологической ниши ареалы. Исключением являются только ареалы родов самых крупных приматов: горилл, орангутанов, мандрил, а также родов очень мелких, но привлекательных для охоты за ними долгопятов.
Единичная, поучительная ситуация возникла при анализе ареала мартышек-гусар. Аналогичная ситуация была с родом отолемуров. Их ареалы не соответствует следствию из принципа Гауза. Проблема решилась, когда на одну карту были нанесены все потенциально конкурирующие за ресурсы саваны рода относительно крупных приматов. Ситуация разобрана на карте саванных приматов Африки.
Рассмотренные фото и видео, в частности, интересны с точки зрения антропоморфности приматов. Особенно она провляется при взаимодействии матерей с младенцами. Часто в этих взаимодействиях степень антропоморфности удивительна. Такие видео мы пометили значком
Предполагаемое развитие проекта
Мы уделяем большое значение картографическому качеству контуров ареалов, карты должны легко читаться (пока это не везде так), контура ареалов на общих границах должны быть качественно прорисованы без наложений. Местами надо уточнять границы ареалов по рекам. Эта картографическая часть работы продолжается.
В работе частично использовались данные получаемые из нейросети Яндекса. В последнее время было замечено, что эта нейросеть улучшила релевантность и часто выдает интересные результаты (с которыми, конечно, надо аккуратно и критически обращаться). Предполагается пропустить все данные презентации через нейросеть, чтобы проверить фактологию и найти новые данные для анализа.
На ФГД пиктограммы современных карт будут заменены на пиктограммы палеокарт.
В работе не использовались данные молекулярной генетики, после получения их от специалистов-приматологов в ФГД будут внесены уточнения. Исправления по результатам консультаций со специалистами-приматологами по ошибкам и неучтенным существенным фактам.
Поиск и оформление лучших мировых фото и видео-материалов по приматам. Презентация должна стать квинтэссенцией всех лучших и самых качественных визуальных материалов по каждому виду приматов. По каждому виду должен быть представлен минимум топ-3 таких материалов.
Гипотеза KF-видообразования
На картах расселения видов отдельных родов приматов, наиболее интересно, кроме почти всегда очевидных случаев аллопатрического и перипатрического видообразования, рассмотреть оставшиеся случаи. Трудно не сделать вывод, что картина расселения видов многих родов приматов часто может быть объяснена только парапатрическим видообразованием.
Причем, это парапатрическое видообразование, кажется, имеет какой-то мощный фактор усиления. Классическим "спокойным" вариантом парапатрического видообразования трудно объяснить, например, карту расселение макак на Сулавеси, больше похожую на политическую карту.
Предложим следующую гипотезу, которую назовем гипотезой KF-видообразования. Кратко смысл такой. Дарвиновская эволюция, почему-то, поддерживает "ген ксенофобии". В результате особи удаленных популяций вида, которые вполне хорошо могут скрещиваться, но уже имеют насколько-то различный фенотип (начальная стадия парапатрического видообразования) начинают скрещиваться меньше, чем могли бы. Предполагаем, что из-за ксенофобии. В результате происходит дробление видов.
Представление проекта как расширения БТК Эволюция в Парке Леко и на сайте проекта Эволюция в каком-то смысле имеет логику обратную методике исследования. Сначала представляется ФГД, затем по каждой части света представлены карты по родам, затем полная картография по каждому семейству обитающему в этой части света с картами по родам и по видам с анализом, сравнительными таблицами и фотогалерея с дополнительными картами и интересными данными по отдельным видам.
Электронный формат предполагает полноценный просмотр в разрешении 4К. В идеале нужно использовать 2 монитора-телевизора 4К.
Зоопарки
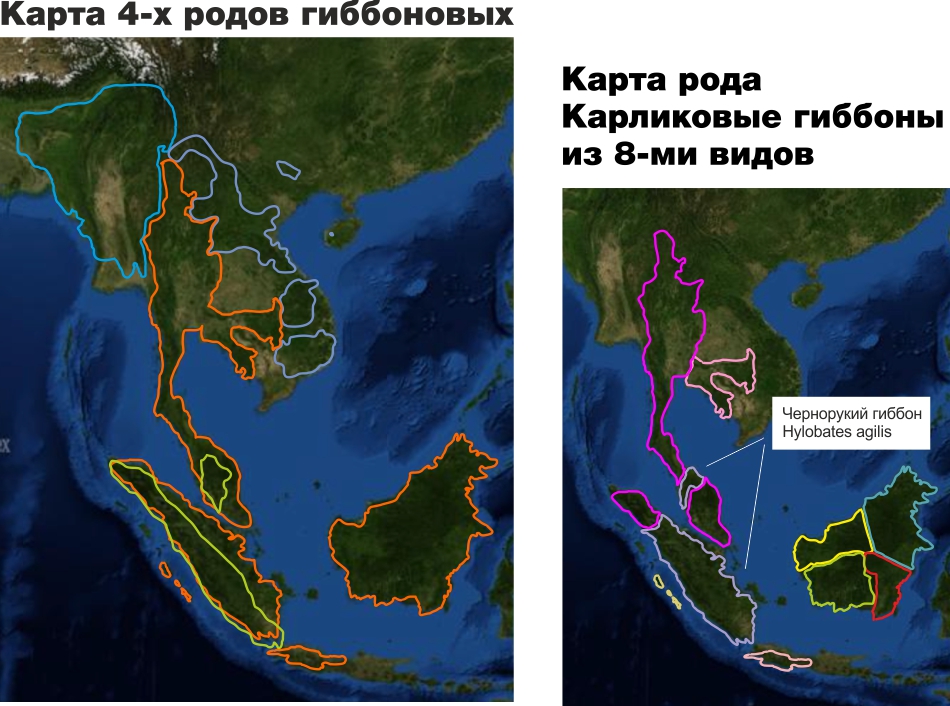
В зоопарках виды всегда представляются примерно так (в дополнение к нижеизложенной критике отметим, что не аккуратный дизайн верхней части в этом конкретном случае не позволяет понять, где официальное наименование вида, включающее официальное название рода, а где неофициальное название вида на английском языке)
Фактически это дезинформация. Неспециалист (не проведя специального самостоятельного исследования в интернете) не поймет, что гиббоны проживают на гораздо более обширной территории, и чернорукий гиббон является одним из 8-ми видов рода карликовых гиббонов, одного из 4-х родов гиббоновых и представляет в Московском зоопарке все семейство гиббоновых.
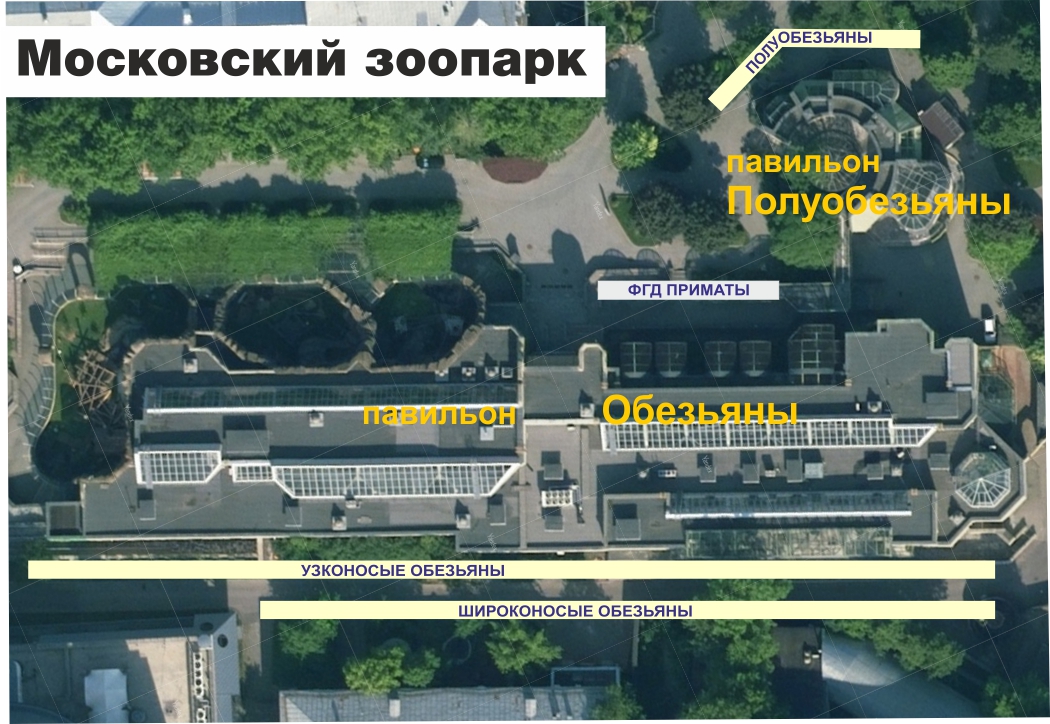
Для экскурсоводов, студентов биофаков, специалистов и всех заинтересованных в изучении зоологии могли бы использоваться такие планшеты
В зоопарках можно было бы выставлять информацию оформленную аналогично нашей презентации. Например, в Московском зоопарке -- вдоль задней аллеи павильона обезьян панели узконосых и широконосых, а полуобезьян рядом с их павильоном.
В идеале такие панели должны быть для всех животных, хотя бы в электронном формате на планшете.
Автор -- Константин Могильницкий Дизайн -- Ольга Кутукова

 ландшафт -- саванна, джунгли, высокогорье и т.д.,
ландшафт -- саванна, джунгли, высокогорье и т.д., питание -- всеядность, насекомоядность, фрукты, орехи, соки растений, листья и т.д.,
питание -- всеядность, насекомоядность, фрукты, орехи, соки растений, листья и т.д., высотный уровень -- поверхность земли, нижний, средний, верхний ярус леса,
высотный уровень -- поверхность земли, нижний, средний, верхний ярус леса, взаимодействие с хищниками,
взаимодействие с хищниками, климатические условия в достаточно широких пределах.
климатические условия в достаточно широких пределах. Аллопатрическое -- географическая изоляция.
Аллопатрическое -- географическая изоляция. Перипатрическое -- частный случай аллопатрического, стратовой ситуацией является изоляция группы в приграничье первоначально общего ареала (важное значение может иметь дрейф генов).
Перипатрическое -- частный случай аллопатрического, стратовой ситуацией является изоляция группы в приграничье первоначально общего ареала (важное значение может иметь дрейф генов). Парапатрическое -- большой ареал имеет некоторые градиенты экологических отличий, который приводит к направленным модификациям вида или существует какой-то ландшафтный рубеж, который преодолим, но с затруднениями или что-то подобное, в результате чего возникает нежесткое разделение на популяции, далее возникают небольшие фенотипические отличия, процесс видообразования завершается под действием фактора избирательности скрещивания.
Парапатрическое -- большой ареал имеет некоторые градиенты экологических отличий, который приводит к направленным модификациям вида или существует какой-то ландшафтный рубеж, который преодолим, но с затруднениями или что-то подобное, в результате чего возникает нежесткое разделение на популяции, далее возникают небольшие фенотипические отличия, процесс видообразования завершается под действием фактора избирательности скрещивания. Гибридизация -- виды с одним кариотипом (числом, размером, формой и т.д. хромосом) даже при существенном различии генотипов, иногда могут дать плодовитое потомство.
Гибридизация -- виды с одним кариотипом (числом, размером, формой и т.д. хромосом) даже при существенном различии генотипов, иногда могут дать плодовитое потомство. Кариотипическое -- одинаковое изменение кариотипа одновременно у мужской и женской особи исключающее возможность давать плодовитое потомство с остальными особями вида, но позволяющее дать плодовитое потомство при скрещивании между собой.
Кариотипическое -- одинаковое изменение кариотипа одновременно у мужской и женской особи исключающее возможность давать плодовитое потомство с остальными особями вида, но позволяющее дать плодовитое потомство при скрещивании между собой. Ароморфическое -- мутация у одной особи дающая существенное преимущество и поддерживаемая естественным или (и) половым отбором (пример -- на филогенетическом дереве приматов -- появление бесхвостых обезьян, ставших позже человекообразными).
Ароморфическое -- мутация у одной особи дающая существенное преимущество и поддерживаемая естественным или (и) половым отбором (пример -- на филогенетическом дереве приматов -- появление бесхвостых обезьян, ставших позже человекообразными). Аллопатрическое, перипатрическое или парапатрическое видообразование состоявшееся достаточно давно, чтобы произошло разделение не только на два вида, но и изменения дошли до столь явных отличий, что виды очень существенно отличаются по ряду признаков.
Аллопатрическое, перипатрическое или парапатрическое видообразование состоявшееся достаточно давно, чтобы произошло разделение не только на два вида, но и изменения дошли до столь явных отличий, что виды очень существенно отличаются по ряду признаков. Симпатрическое видообразование состоявшееся достаточно давно. Но с позиций "экологического" определения рода ситуация должна быть описана по другому. Момент состоявшегося симпатрического видообразования нужно считать моментом разделения рода на два. Т.е. образование двух видов и двух родов при симпатрическом видообразовании -- одно и тоже событие.
Симпатрическое видообразование состоявшееся достаточно давно. Но с позиций "экологического" определения рода ситуация должна быть описана по другому. Момент состоявшегося симпатрического видообразования нужно считать моментом разделения рода на два. Т.е. образование двух видов и двух родов при симпатрическом видообразовании -- одно и тоже событие. Мгновенное видообразование ("кариотипическое" и "араморфическое") -- новый вид (уже сразу род) выживет, если покинет ареал старого вида или вытеснит старый вид.
Мгновенное видообразование ("кариотипическое" и "араморфическое") -- новый вид (уже сразу род) выживет, если покинет ареал старого вида или вытеснит старый вид. Симпатрическое ответвление -- ответвление с наклоном
Симпатрическое ответвление -- ответвление с наклоном 
 Стабилизирующая замена -- прямая со сменяющими друг друга без ответвлений родами
Стабилизирующая замена -- прямая со сменяющими друг друга без ответвлений родами
 Аллопатрическое ответвление (миграция) -- вертикальное ответвление
Аллопатрическое ответвление (миграция) -- вертикальное ответвление
 Аллопатрическое разветвление -- вертикальные вверх и вниз ответвления
Аллопатрическое разветвление -- вертикальные вверх и вниз ответвления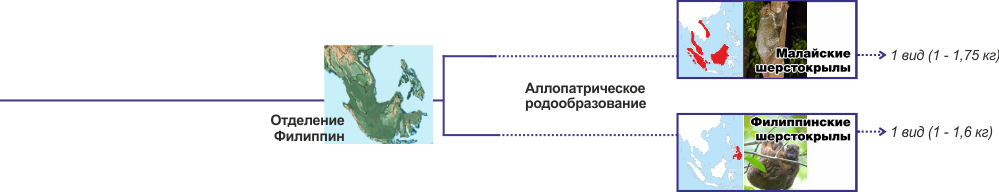
 Парапатрическое родообразование -- расходящиеся наклонные от карты предкового рода к картам новых родов
Парапатрическое родообразование -- расходящиеся наклонные от карты предкового рода к картам новых родов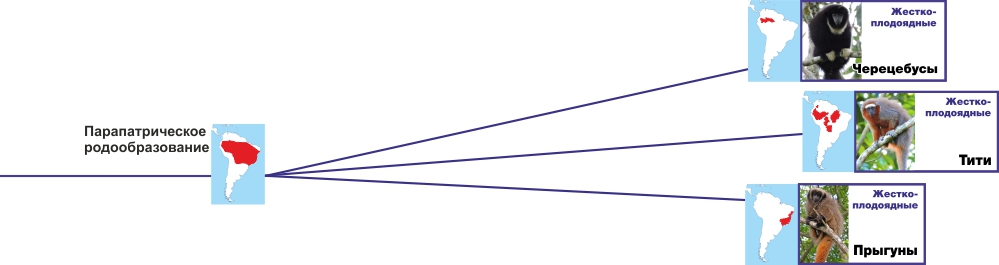
 Ароморфическое ответвление -- вертикальное ответвление
Ароморфическое ответвление -- вертикальное ответвление
 Вымирание -- крестик
Вымирание -- крестик 
 Современное незаконченное событие родообразования -- в рамке показыается не род, а вид, используются зеленые линии. Подпись: "Текущий процесс родообразования".
Современное незаконченное событие родообразования -- в рамке показыается не род, а вид, используются зеленые линии. Подпись: "Текущий процесс родообразования". 

 Удивительно, что столь простой, использованный нами, метод исследования является в основном достаточным для определения экологических ниш, что свидетельствует об ограниченном их количестве и о полноте их занятия. Нет экологически возможных и не занятых ниш. Одни и те же ниши в разных частях света заняты самыми разными и филогенетически очень далекими родами приматов.
Удивительно, что столь простой, использованный нами, метод исследования является в основном достаточным для определения экологических ниш, что свидетельствует об ограниченном их количестве и о полноте их занятия. Нет экологически возможных и не занятых ниш. Одни и те же ниши в разных частях света заняты самыми разными и филогенетически очень далекими родами приматов. Экологических ниш довольно небольшое и конкретное количество. Ниши более обширны, чем можно было предполагать, например, не нашло подтверждения предположение, что разные рода листоядных в одном ареале питаются листьями разных деревьев, такого относительно узкого разделения ниш нет. Практически нет случаев, где бы экологическая ниша определялась хищниками (исключением, вероятно, являются черно-белые и красные колобусы). Экологические ниши определяются конкуренцией за пищу, т.е. являются почти строго пищевыми.
Экологических ниш довольно небольшое и конкретное количество. Ниши более обширны, чем можно было предполагать, например, не нашло подтверждения предположение, что разные рода листоядных в одном ареале питаются листьями разных деревьев, такого относительно узкого разделения ниш нет. Практически нет случаев, где бы экологическая ниша определялась хищниками (исключением, вероятно, являются черно-белые и красные колобусы). Экологические ниши определяются конкуренцией за пищу, т.е. являются почти строго пищевыми. Почти не обнаружено случаев несоответствия принципу Гауза при анализе видов одного рода.
Почти не обнаружено случаев несоответствия принципу Гауза при анализе видов одного рода. Следствие из принципа Гауза нарушается только для родов: мандрилы, гелады, мартышки-гусары, отолемуры. В каждом случае приведено объяснение (если нам удалось найти объяснение в источниках) или высказано предположение об этом несоответствии (если мы не нашли объяснения в источниках).
Следствие из принципа Гауза нарушается только для родов: мандрилы, гелады, мартышки-гусары, отолемуры. В каждом случае приведено объяснение (если нам удалось найти объяснение в источниках) или высказано предположение об этом несоответствии (если мы не нашли объяснения в источниках). Антропогенное воздействие на приматов на уровне видов очень существенно, однако при взгляде на уровне родов нарушение следствия из принципа Гауза незначительно. Рода заселяют практически все соотвествующие их экологической ниши ареалы. Исключением являются только ареалы родов самых крупных приматов: горилл, орангутанов, мандрил, а также родов очень мелких, но привлекательных для охоты за ними долгопятов.
Антропогенное воздействие на приматов на уровне видов очень существенно, однако при взгляде на уровне родов нарушение следствия из принципа Гауза незначительно. Рода заселяют практически все соотвествующие их экологической ниши ареалы. Исключением являются только ареалы родов самых крупных приматов: горилл, орангутанов, мандрил, а также родов очень мелких, но привлекательных для охоты за ними долгопятов. Единичная, поучительная ситуация возникла при анализе ареала мартышек-гусар. Аналогичная ситуация была с родом отолемуров. Их ареалы не соответствует следствию из принципа Гауза. Проблема решилась, когда на одну карту были нанесены все потенциально конкурирующие за ресурсы саваны рода относительно крупных приматов. Ситуация разобрана на карте саванных приматов Африки.
Единичная, поучительная ситуация возникла при анализе ареала мартышек-гусар. Аналогичная ситуация была с родом отолемуров. Их ареалы не соответствует следствию из принципа Гауза. Проблема решилась, когда на одну карту были нанесены все потенциально конкурирующие за ресурсы саваны рода относительно крупных приматов. Ситуация разобрана на карте саванных приматов Африки. Рассмотренные фото и видео, в частности, интересны с точки зрения антропоморфности приматов. Особенно она провляется при взаимодействии матерей с младенцами. Часто в этих взаимодействиях степень антропоморфности удивительна. Такие видео мы пометили значком
Рассмотренные фото и видео, в частности, интересны с точки зрения антропоморфности приматов. Особенно она провляется при взаимодействии матерей с младенцами. Часто в этих взаимодействиях степень антропоморфности удивительна. Такие видео мы пометили значком

 Мы уделяем большое значение картографическому качеству контуров ареалов, карты должны легко читаться (пока это не везде так), контура ареалов на общих границах должны быть качественно прорисованы без наложений. Местами надо уточнять границы ареалов по рекам. Эта картографическая часть работы продолжается.
Мы уделяем большое значение картографическому качеству контуров ареалов, карты должны легко читаться (пока это не везде так), контура ареалов на общих границах должны быть качественно прорисованы без наложений. Местами надо уточнять границы ареалов по рекам. Эта картографическая часть работы продолжается. На ФГД пиктограммы современных карт будут заменены на пиктограммы палеокарт.
На ФГД пиктограммы современных карт будут заменены на пиктограммы палеокарт. Исправления по результатам консультаций со специалистами-приматологами по ошибкам и неучтенным существенным фактам.
Исправления по результатам консультаций со специалистами-приматологами по ошибкам и неучтенным существенным фактам. Поиск и оформление лучших мировых фото и видео-материалов по приматам. Презентация должна стать квинтэссенцией всех лучших и самых качественных визуальных материалов по каждому виду приматов. По каждому виду должен быть представлен минимум топ-3 таких материалов.
Поиск и оформление лучших мировых фото и видео-материалов по приматам. Презентация должна стать квинтэссенцией всех лучших и самых качественных визуальных материалов по каждому виду приматов. По каждому виду должен быть представлен минимум топ-3 таких материалов.
 Представление проекта как расширения БТК Эволюция в Парке Леко и на сайте проекта Эволюция в каком-то смысле имеет логику обратную методике исследования. Сначала представляется ФГД, затем по каждой части света представлены карты по родам, затем полная картография по каждому семейству обитающему в этой части света с картами по родам и по видам с анализом, сравнительными таблицами и фотогалерея с дополнительными картами и интересными данными по отдельным видам.
Представление проекта как расширения БТК Эволюция в Парке Леко и на сайте проекта Эволюция в каком-то смысле имеет логику обратную методике исследования. Сначала представляется ФГД, затем по каждой части света представлены карты по родам, затем полная картография по каждому семейству обитающему в этой части света с картами по родам и по видам с анализом, сравнительными таблицами и фотогалерея с дополнительными картами и интересными данными по отдельным видам. Электронный формат предполагает полноценный просмотр в разрешении 4К. В идеале нужно использовать 2 монитора-телевизора 4К.
Электронный формат предполагает полноценный просмотр в разрешении 4К. В идеале нужно использовать 2 монитора-телевизора 4К.