|
|
Цвета шрифта в тексте обозначают:
Черный -- общепринятые научные данные или формулировки автора, которые требуют научной редакции, но вряд ли сильно искажают общепринятые смыслы, а так же изложение гипотезы содержащее простые логические построения.
Зеленый -- тезисы и теоретические модели имеющие целью сделать текст логически связанным, они использованы в случаях, когда автор курса не нашел ясных общепринятых объяснений, а так же другие оригинальные части текста.
Оранжевый -- аналог зеленого, очень спорные, но внутренне не противоречивые версии и рассуждения, а так же непроверенные нормальным образом тезисы, высказываемые автором в утвердительной форме.
Гипотеза генезиса нуклеотидов на основе предшествующих нуклеоноидов
+ предположение о неферментативном синтезе РНК на основе нуклеоноидов
Вся биологическая информация записывается в виде полимера РНК или ДНК.
Алфавитом для этой записи являются 4 молекулы -- нуклеотиды.
Для РНК они выглядят так:
А Г Ц У
Эти молекулы создаются живыми организмами. Почему именно конкретно эти молекулы, изначально возникшие абиогенно, являются алфавитом для записи биологической информации, не известно. Работы в этом направлении ведутся, но сводятся к правдоподобным предположениям о предшественниках нуклеотидов и поиску их в экспериментах аналогичных экспериментам проводимым лабораторией Джона Сазерленда для самих нуклеотидов.
Ниже представлена гипотеза объясняющая генезис нуклеотидов через предшествующих им нуклеоноидов, а так же, как следствие предположение об успешном неферментативном синтезе РНК на основе предшествующих нуклеотидам нуклеоноидов.
Гипотеза пока не прошла научного рецензирования.
Гипотеза демонстрирует возможный путь объяснения причин возникновения 4 млрд. лет назад именно АУЦГ-алфавита РНК. Это знание должно быть достигнуто, оно является фундаментальным для биологии, аналогично нукеосинтезу для физики.
Граница трех сред
В результате геологической эволюции, источником энергии для которой является гравитационная дифференциация, через несколько десятков миллионов лет поверхность планеты становится океаном усеянном вулканическими островами.

Возникает новая среда -- граница трех сред. Эта граница существует на нескольких видах локаций: дождевые лужи, лужи на побережьях, гейзерные лужи, грязевые котлы у вулканов.
Периодическое высыхание луж и подсыхание грязевых котлов на планете с водой в жидкой форме позволяет не только создавать более высокие концентрации мономеров, но и смещает равновесие между созданием цепочки и ее растворением в пользу полимеризации, удаляя избыток воды.

На границе трех сред при ослабленном действии растворителя могут возникать новые виды молекул -- супрамолекулы и супраполимеры.
Поляризационные связи между азотистыми гетероциклами
Возникновение межмолекулярного поляризационного водородного взаимодействия имеет те же причины, из-за которых вещества являются хорошими растворителями. Поляризованные концы молекул притягиваются друг к другу и к частям других молекул обладающих таким же свойством. Возникают связи между атомами -- межмолекулярные связи. Они намного слабее внутримолекулярных ковалентных связей, но для эволюции играют решающую роль.
Далее рассматриваются межмолекулярные связи между гетероциклами. Опускаются рассуждения почему все другие виды молекул не годятся для целей создания алфавита жизни. Достаточным основанием считаем, что именно на гетероциклах построен действующий алфавит.
Межмолекулярные связи через водород создаются между атомами фтора, кислорода и азота. Водородные связи через фтор самые сильные, но фтора в природе мало, на втором месте по силе водородные связи через кислород и самые слабые через азот. Таким образом, водородные межмолекулярные связи имеющие существенное значение создаются только атомами кислорода и азота.

Есть химическая причина (ее надо прописать, она точно есть) по которой азотистые гетероциклы не могут содержать гидроксильных групп (-OH) и не могут создавать самой сильной поляризационной связи, т.е. поляризационные связи между азотистыми гетероциклами могут создаваться только тремя способами.
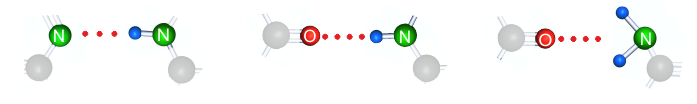
Чтобы между циклическими молекулами образовалась хотя бы одна поляризационная связь, они должны содержать в контуре цикла минимум один атом азота, т.е. быть азотистыми гетероциклами.
Супрамолекулы с тройными связями
Чем больше поляризационных связей, тем прочнее гетероциклы связаны и тем стабильнее возникающая конструкция, и в условиях границы трех сред при ослабленном влиянии растворителя она может существовать достаточно долгое время. Пары молекул связанных поляризационными связями называются супрамолекулами.
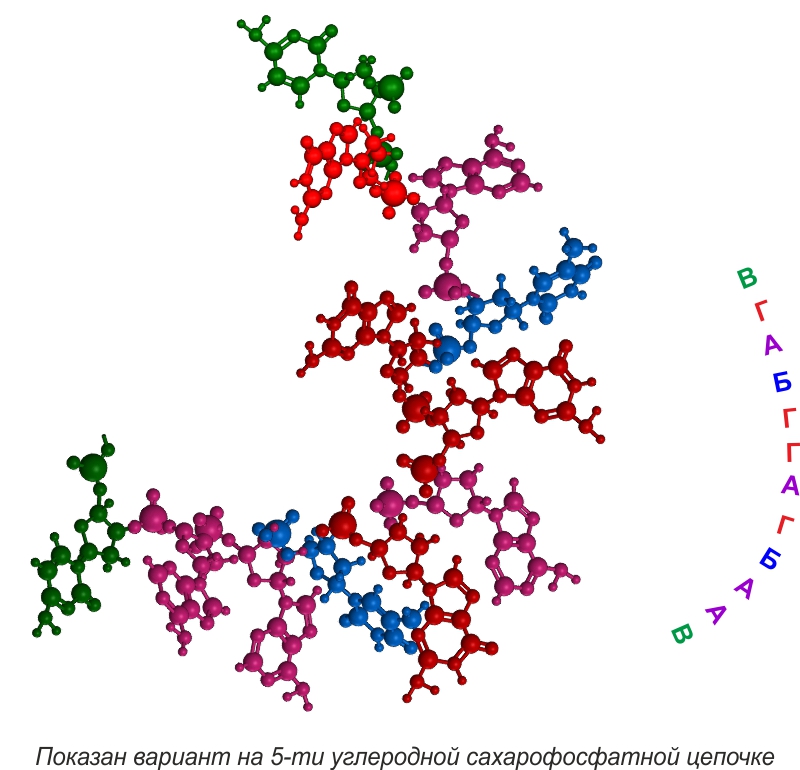
Введем условное обозначение для 5-ти и 6-ти атомных гетероциклов. 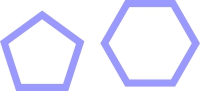
5-ти угольные циклы геометрически могут создать супрамолекулы только с двумя связями, 6-ти угольные геометрически могут создавать супрамолекулы с тремя связями.
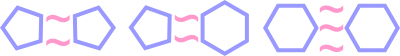
Введем условие №1: для прочной стабильной связи гетероциклы должны быть связаны тремя поляризационными связями.
Для 6-ти угольных гетероциклов существует всего 8 возможных комбинаций с поляризационными связями.
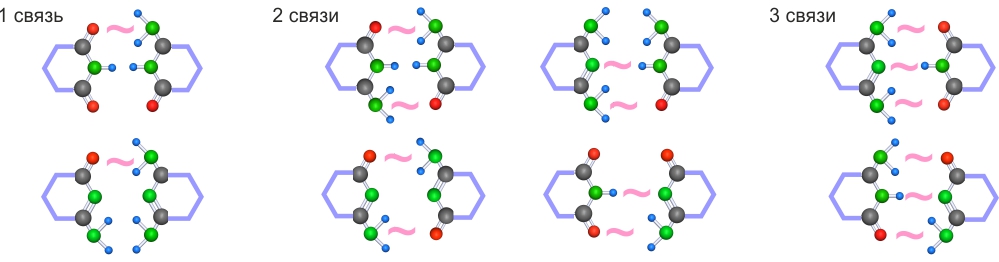
Из 8-ми возможных комбинаций на атомах, которые геометрически могут создавать поляризационные связи, только 2 пары гетероциклов соответствуют условию №1 -- могут создавать между собой тройные связи (без учета вариантов образуемых на трех атомах, которые геометрически не могут участвовать в поляризационных связях).
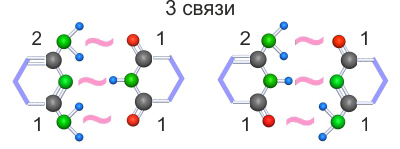
Назовем сторону, которая участвует в трех поляризационных связях, поляризационной стороной. Заметим, что в каждой супрамолекуле с тремя поляризационными связями один гетероцикл имеет между поляризационной и неполяризационной сторонами конфигурацию ковалентных связей 2/1, а второй 1/1.
В общем виде супрамолекулы с тройными связями выглядят так.
Супрамолекулы устойчивые к растворению
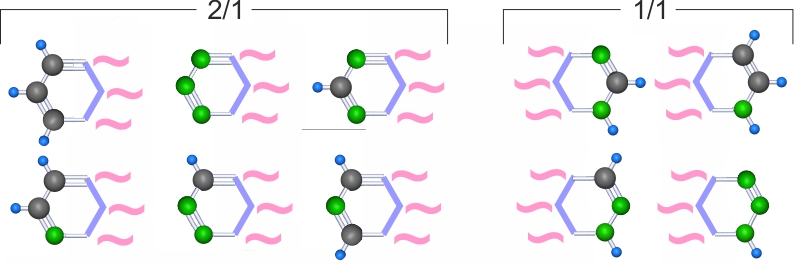
Для противодействия растворению в водном растворе на неполяризационной стороне каждого гетероцикла в паре должно быть не более одного слабого поляризационного конца. Назовем это условием №2 .
Для конфигурации 2/1 с учетом условия №2 возможны 6 вариантов, для конфигурации 1/1 -- 4 варианта.
Способность к супраполимеризации
Следующий эволюционный уровень -- объединение супрамолекул в полимеры. Для этого супрамолекулам необходимо объединиться с молекулами способными создавать полимеры, это сахорофосфаты и аминокислоты.
Присоединение к сахарофосфатам или аминокислотам происходит через слабую поляризационную связь.
Супрамолекулы могут ковалентно присоединять сахарофосфаты или аминокислоты, но для этого они должны иметь на неполяризационной стороне именно один слабый поляризационный конец, через который произойдет присоединение. Назовем этот более строгий вариант условия №2 условием №3 . Конфигурации 1/1 соответствуют условию №3, а конфигурации 2/1 не соответствуют. Для несоответствующих условию №3 пиримидинов условию №3 соответствуют производные от них пурины, но соединение 6-ти угольного и 5-ти угольного гетероцикла может происходить только через два соседних атома углерода.

Пурины могут создаваться только на двух пиримидинах конфигурации 2/1.
Для каждого из этих случаев существует 2 варианта пурина соответствующих условию №3.
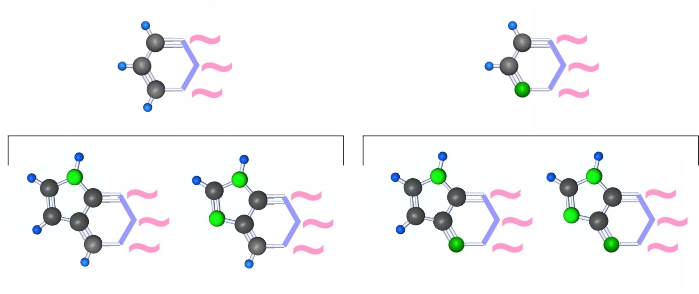
Общее число способных к полимеризации супрамолекул (теперь пиримидино-пуриновых) составляет 32.

Супраполимеры
Удовлетворяющие условиям №1 и №3 пиримидин-пуриновые супрамолекулы, присоединив аминокислоты или сахарофосфаты, могли создавать супрамономеры защищенные от растворителя и способные к полимеризации.
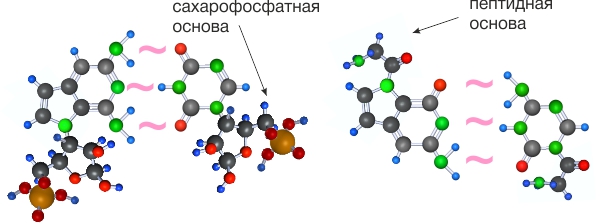
Супрамономеры объединяются в супраолигомеры.
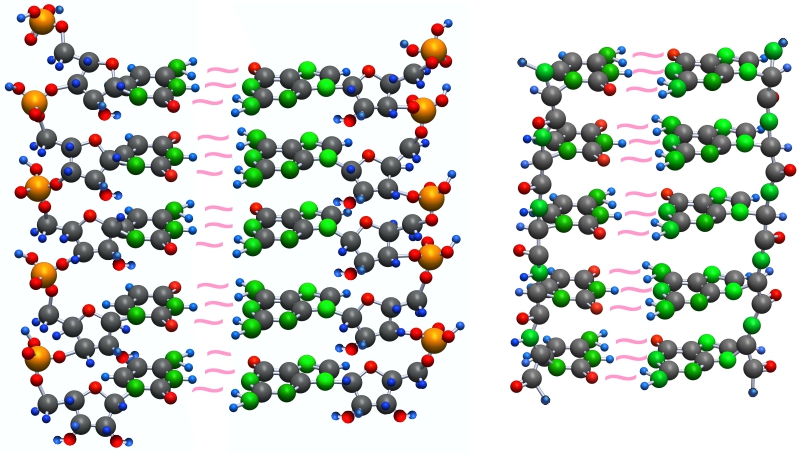
При супраполимеризации такие цепочки образуют спирали.
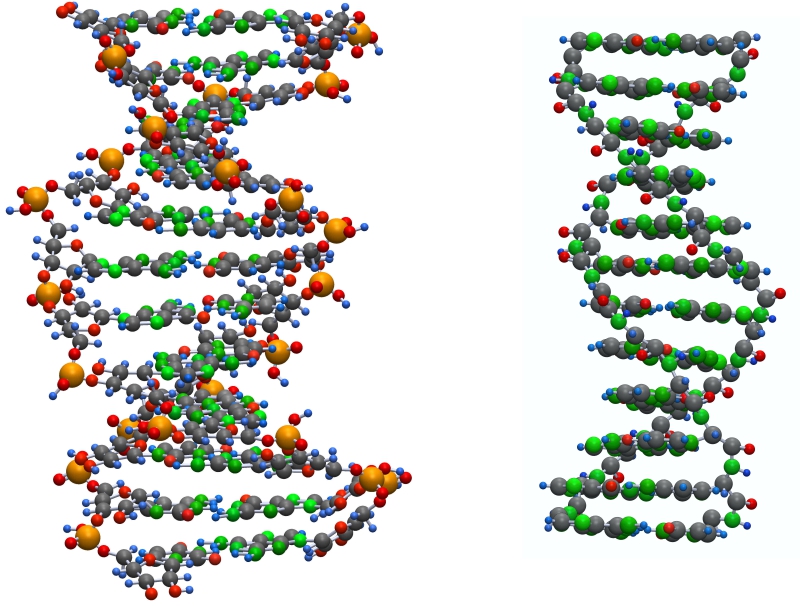
Температурный фактор
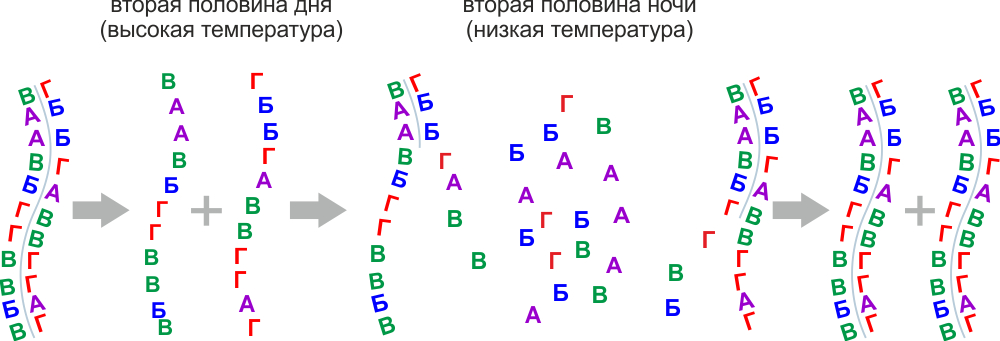
При нагревании супраполимеры распадаются сначала по поляризационным связям, образуя однониточные цепочки, и далее на мономеры трехчленные для сахарофосфатных и двучленные для пептидных полимеров.
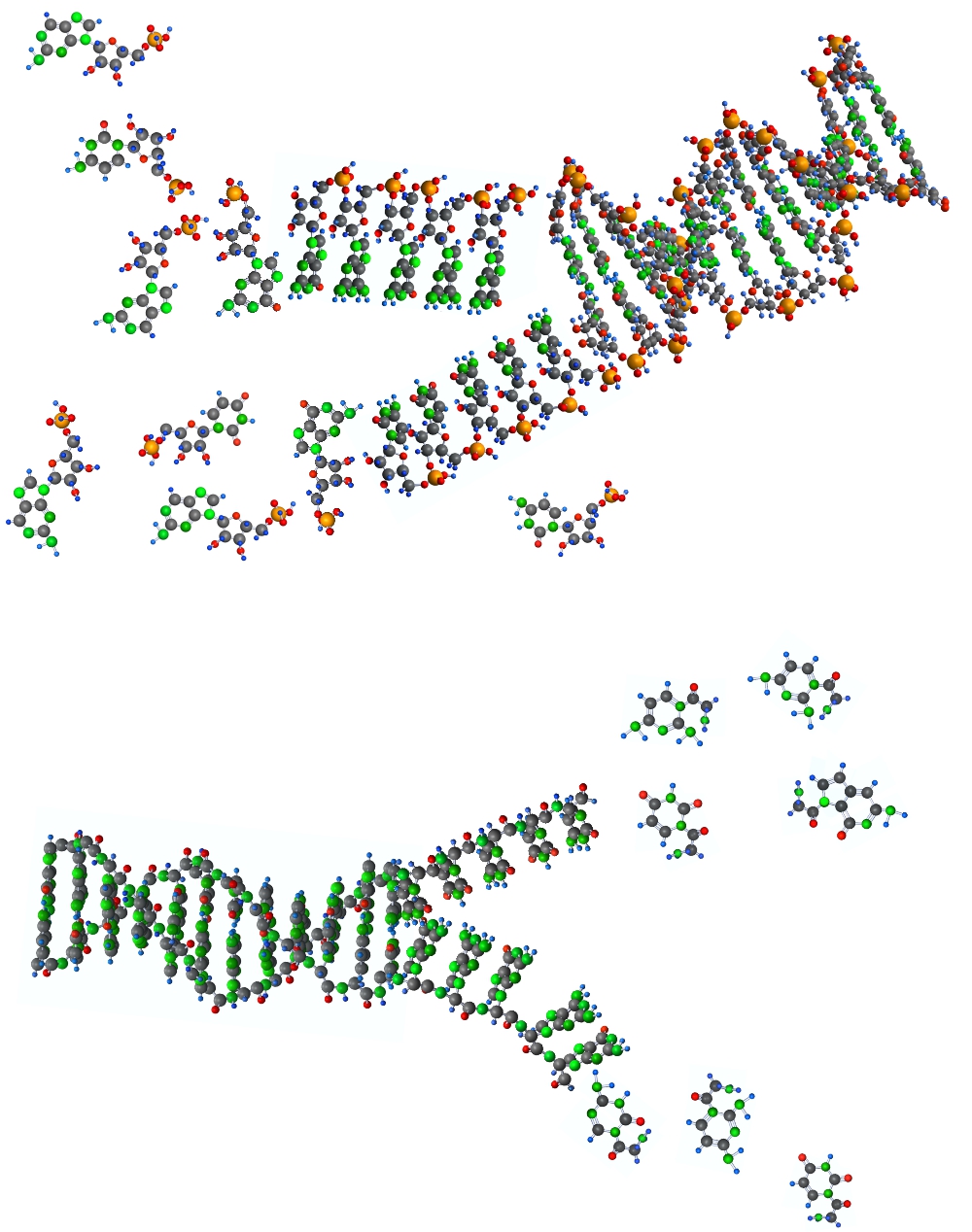
Дальнейшая полимеризация и супраполимеризация идет уже между такими мономерами.
|
|
Супраполимеризация, УФ-отбор и пояс жизни
Сахарофосфатные и пептидные мономеры способные к супраполимеризации из-за обилия вариантов не могли создать нового эволюционного качества, нужен некий отбор, уменьшающий количество вариантов.
Диазотная антена, нуклеоноиды
По некоторым причинам, которые изучает квантовая механика или которые следуют из экспериментов (у меня это просто результат анализа, чем отличаются нуклеотидные супрасвязанные азотистые основания способные к полимеризации от других из этих 32), устойчивы к УФ-облучению только гетероциклы с двумя атомами азота .
Из 32-х супрамолекул способных к полимеризации только 2 имеют по 2 атома азота в каждом гетероцикле, т.е. из 32-х супрамолекул УФ-облучение выжигает 30, остаются только 2. В результате, в условиях УФ-облучения устойчивые супрамолекулярные полимеры могут создаваться только двумя УФ-устойчивыми парами.
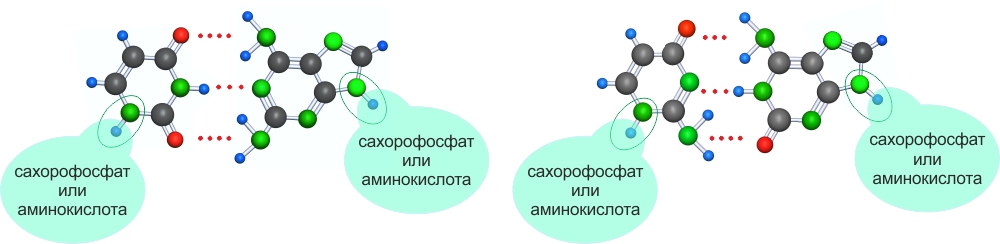
Дополнительно ("неожиданно", это не следовало из предыдущих рассуждений), в результате отбора на растворение, отбора на способность связываться с мономерами способными к полимеризации и УФ-отбора возникло новое качество -- однозначность (каждой из 4-х молекул соответствует в качестве партнера по паре только одна). Однозначность является необходимым свойством для возникновения нового качества полимеров, полимеры строящиеся на однозначных мономерах могут сохранять и размножать информацию. Назовем 4 вида молекул способных создавать однозначные пары нуклеоноидами.
Однозначность нуклеоноидовых супрамолекул позволила эволюции перейти на новый уровень.
Далее будут рассматриваться только нуклеоноиды, их полимеризация и супраполимеризация.
Супраполимеризация УФ-устойчивых нуклеоноидов возможна только на планетах, находящихся на расстоянии от 100 до 250 млн. км от звезды, где достаточно сильное УФ-облучение разрушающее конкурирующие с нуклеоноидами азотистые гетороциклы сосуществует с границей трех сред.
|
|
Нуклеоноиды
Итак, на основе сахарных и пептидных цепочек возникают мономеры с 4-мя разными азотистыми гетероциклами -- нуклеоноиды.
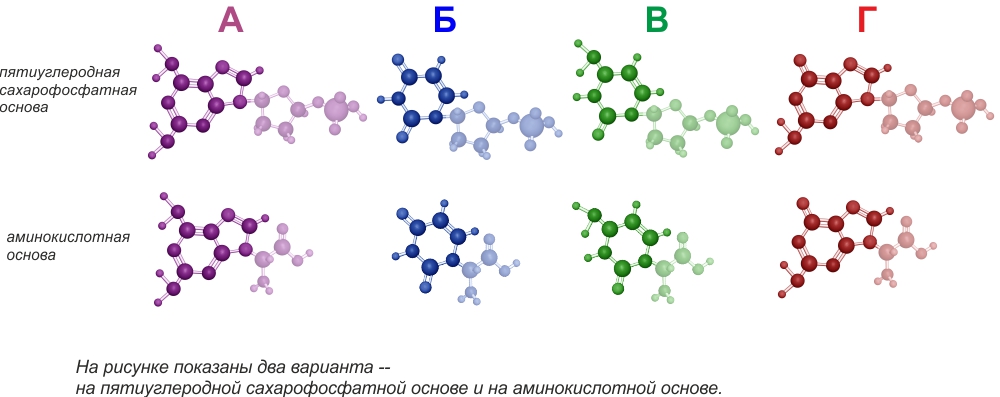
Для дальнейшего удобства 4 мономера обозначены цветом и буквами А, Б, В, Г.
Полимеризация
При понижении температуры сшивались уже мономеры – готовые звенья цепи. Сшивание могло быть просто в одиночную цепочку (обычно на какой-то минеральной подложке).
Репликация
(здесь и в нескольких параграфах ниже информация, на первый взгляд, уровня школьного учебника, но это потому, что трудно отделить эту информацию от информации содержащей новизну метода объяснения и приводящую к мысли о неферментативном синтезе РНК на основе нуклеоноидов)
Сшивание могло происходить в виде строительства цепи "с нуля" или супрамолекулярным способом на уже существующей (сохранившейся) одиночной цепи. В результате получалась двойная цепь. Этот способ сшивания называется репликацией.
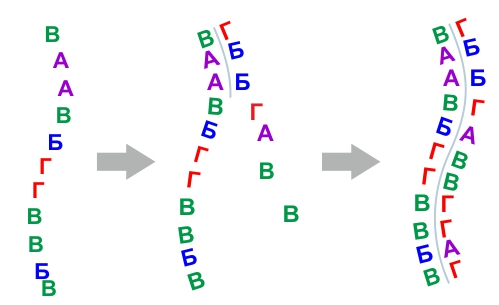
Так же, если двойная цепь распалась на две, и они обе не распались далее, то строительство происходило на каждой из них, это тоже репликация.
Однозначность -- основа наследственности
Важнейшая особенность нуклеоноидов -- создавать однозначные пары: А создает пару только с Б, а В создает пару только с Г.
Однозначность, наблюдаемая в этой закономерности, эквивалентна однозначности в информационной цепочке, создаваемой таким основным информационным инструментом, как алфавит. Информация, записанная с помощью алфавита, может быть скопирована и размножена, т.е. переходить без потери содержания из поколения в поколение. В данном случае алфавит 4-х буквенный.
Из-за однозначности при репликации новая двойная цепь получалась одинаковой с первоначально распавшейся, причем независимо от того, на какой из двух одиночных цепей происходило восстановление. Это создало механизм наследственности.
Биохимия
Мономеры способные реплицироваться называются биомономерами.
Репликация предопределила переход эволюции с химического уровня на биохимический (уровень сложности, при котором процессы не могут быть исследованы в пределах химической науки, т.к. начинают действовать естественные информационные системы, подчиняющиеся своим законам).
Проблема выживаемости реплицирующихся цепочек
Цепочки на пептидной основе (из-за отсутствия распределенного электрического заряда, который имеется на сахорофосфатной основе) сворачивались слишком сильно, плохо распрямлялись и служили плохой матрицей для репликации. Они не могли реплицироваться на достаточно длинных участках и не смогли составить конкуренцию цепочкам на сахарофосфатной основе. Из-за этого в среде преимущественно накапливались трехкомпонентные нуклеоноиды на основе 3-, 4-х и 5-ти углеродных сахорофосфатов.
Цепь могла существовать несколько поколений, размножаться, проходя через расшивание цепей и новое строительство, пока не разрушалась в результате ошибок и под действием внешних факторов разрушающих обе цепи.
Эволюция смогла перейти с химического на биохимический уровень сложности не только вследствие однозначности при репликации, но и благодаря еще двум химическим свойствам реплицирующихся цепочек биомономеров с азотистыми гетероциклами -- изменчивость и химическая разница комплиментарных цепочек.
Частота изменчивости и сила химической разности комплементарных цепочек оказались точно подходящими для жизнеспособности реплицирующихся цепочек. Невозможно представить дальнейшую эволюцию при сколько-нибудь существенном нарушении количественных характеристик хотя бы одного из этих свойств. Мы еще раз можем сказать: Почему это так мы не знаем, так устроена наша Вселенная. Подробнее здесь.
Изменчивость -- предпосылка разнообразия
Водородные межмолекулярные связи между нукленоидами допускали редкие возможности ошибок при копировании. Такая ошибка при нормальных условиях спонтанно возникает приблизительно одна на пятьдесят актов встраивания нуклеоноида в последовательность строящуюся на матричной цепи.
Способность к созданию двойных спиралей с не слишком малым и с не слишком большим количеством ошибок (замен) и без существенного нарушения прочности двойной спирали явилась важнейшим химическим свойством нуклеоноидов.
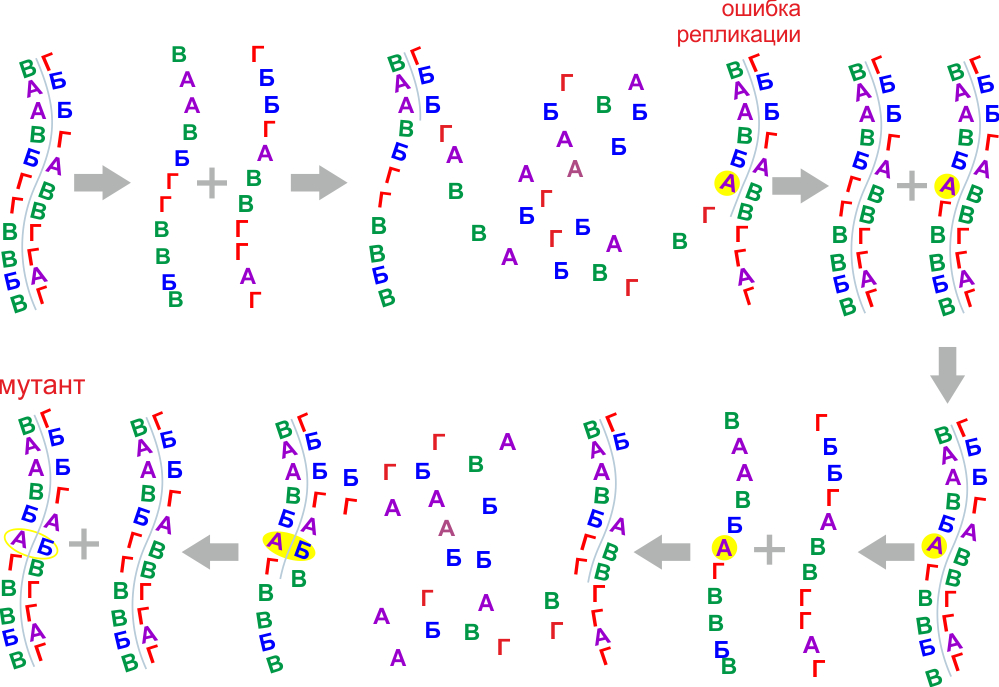
Случайная замена нуклеоноида в процессе репликации (ошибка) называется мутацией. Механизм репликации сразу же закрепляет мутацию, и в последующих поколениях мутация является полноправной частью наследственной информации.
Таким образом, появляется необходимое для эволюции свойство -- изменчивость, позволяющая генерировать новые комбинации на основе старых с единичными заменами. Начался наследственный отбор, дольше выживали более успешные цепочки (являющиеся случайным результатом проявления свойства изменчивости для биомономеров) и на локации происходило накопление успешных обрывков цепей. Это наследственная эволюция.
Разность химических свойств комплементарных цепочек -- предпосылка структурной каталитической активности
Тиражирование информации записанной с помощью алфавита может быть прямым (как в настоящем алфавите), когда букве А соответствует буква А, букве Б буква Б, В - В, Г - Г. Но можно вообразить искусственный алфавит, где соответствие между буквами может быть взаимно-однозначным, но не прямым, когда букве А всегда соответствует буква Б и наоборот, а букве В буква Г и наоборот, а слова записанные таким образом имеют одинаковое значение. Например, слово АААБА имеет такой же смысл, что и слово БББАБ. Второй способ считывания и тиражирования информации называется комплементарным.
При распаде двойной спирали, цепочки на 3-х, 4-х и 5-ти углеродном сахаре могла происходить репликация, в результате которой появлялись клоны молекул, но в результате низкой скорости процесса и низкой точности репликации, через несколько поколений происходило разрушение этого процесса. Однако благодаря комплементарности существует возможность, которая может повысить скорость размножения.
Поскольку информация записана с помощью химических мономеров, то, несмотря на то, что комплементарные цепочки информационно одинаковы, химически они не одинаковы. При распаде двойной спирали цепочки, имея комплементарную последовательность нуклеоноидов и служащие шаблоном для восстановления двух одинаковых двойных спиралей, т.е. несущие одинаковую наследственную информацию, тем не менее обладают разными химическими свойствами. Некоторые куски цепочек, имеющих удачную последовательность нуклеоноидов, могли свернуться в некоторую конфигурацию, когда комплементарный кусок этой цепочки продолжал существовать в несвернутом виде.

На достаточно длинных полимерных цепочках способных к сворачиванию впервые возникла новая функциональная химическая возможность, обусловившая дальнейшую эволюцию. Некоторые из этих конфигураций строго специализированно воздействовали на окружающие молекулы (являясь функциональными аналогами минеральных поверхностей, оказывающих структурное каталитического воздействие на химические реакции). За счет создания карманов, ложбин и разных геометрических объемных структур, удачные варианты способствовали повышению скорости прохождения конкретных реакций, т.к. могли очень точно ориентировать молекулы (включая сложные и крупные), вступающие в реакции относительно друг друга.
Свернувшаяся цепь со временем разрушалась, но это время могло многократно превышать время цикла репликации. И иногда случайно возникавшая конфигурация обладала свойством так влиять на взаимодействие окружающих молекул, что прямо (например, способствуя сшиванию обрывков цепи) или косвенно (например, обогащая среду нужными для репликации молекулами) улучшала способность к размножению своего комплементарного двойника, т.е. способствовала повышению скорости своей же репликации.
Рибозные нуклеоноиды. Рибозимы
Разное химической поведение комплементарных цепочек сильнее проявлялась на цепочках построенных на 5-ти углеродном (рибозном) компоненте, имеющем активные группы -ОН. Цепочки на 3-х и 4-х углеродном сахаре, имеющие такую же разность в составе, были более инертны к этой разнице, она у них проявлялась слабее, т.к. сворачивание без группы -ОН возможно, но происходит намного реже.
Полимеризованные нуклеоноиды на основе рибозы, обладающие свойствами ускорять с помощью механизма структурного каталитического воздействия конкретную химическую или биохимическую реакцию, называют рибозимами .
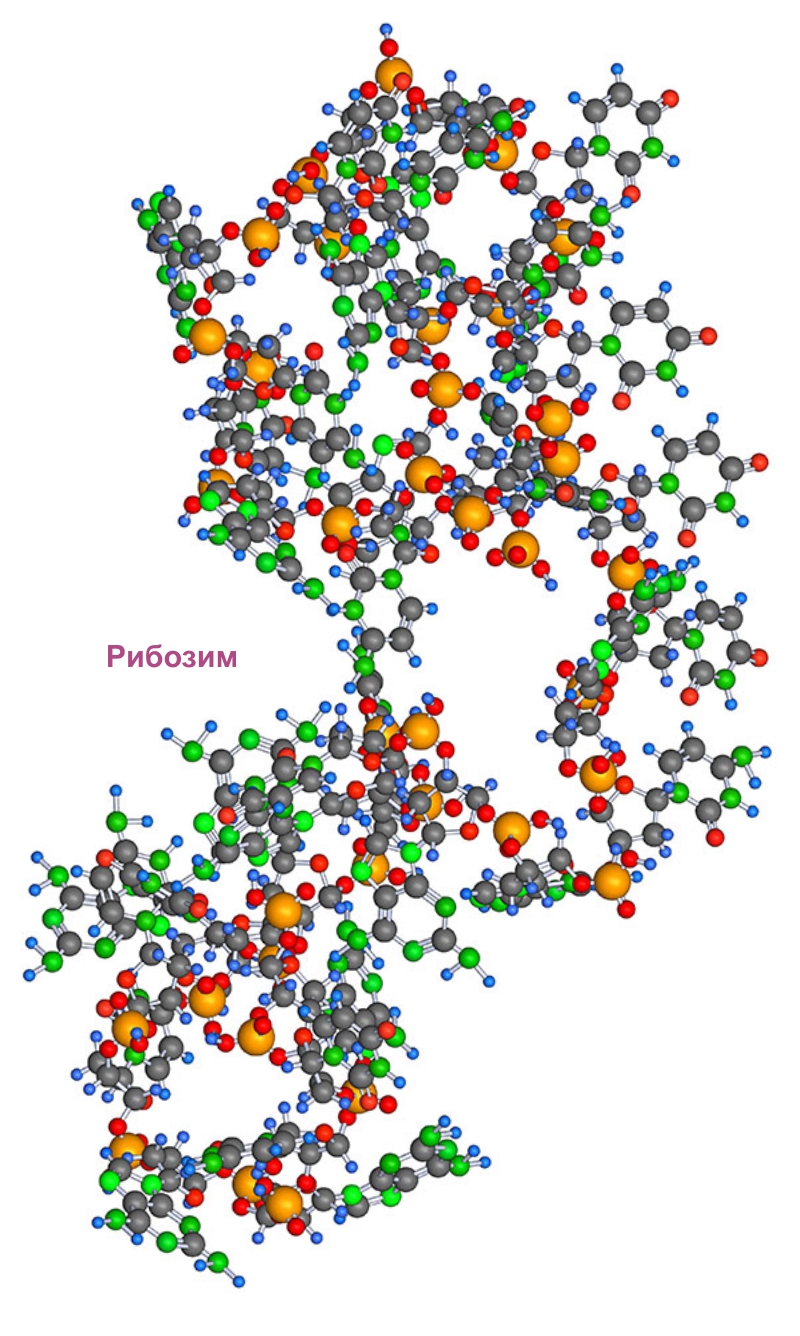
|
|
Нуклеотиды
Еще лучше рибозимные свойства проявились, когда устойчивость цепочек за счет действия рибозимов повысилась настолько, что прочность водородных связей между комплементарными нуклеоноидами могла несколько ослабнуть. В результате изменения в молекуле А в одной паре тройная водородная связь заменилась на двойную.
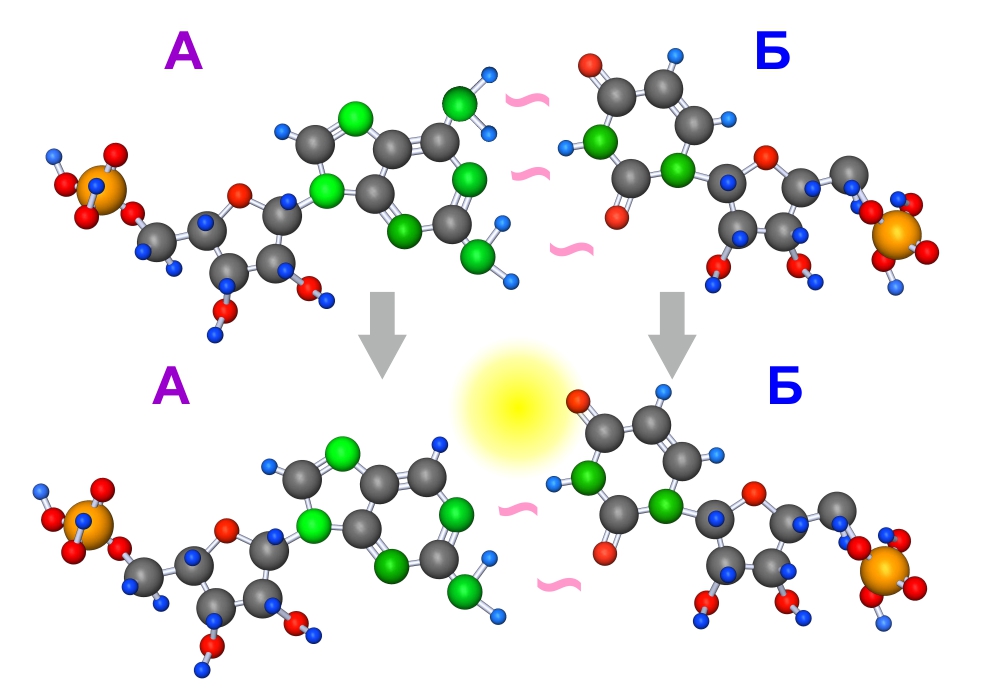
Ухудшение прочности межмолекулярного взаимодействия компенсировалось повышением способности создавать более сложные и разнообразные пространственные конфигурации, т.е. способствовало эволюции рибозимов.
Эти четыре азотисто-рибозно-фосфатные молекулы называются нуклеотидами (из-за того, что впервые были обнаружены в клеточном ядре, которое в английском языке называется nucleus).
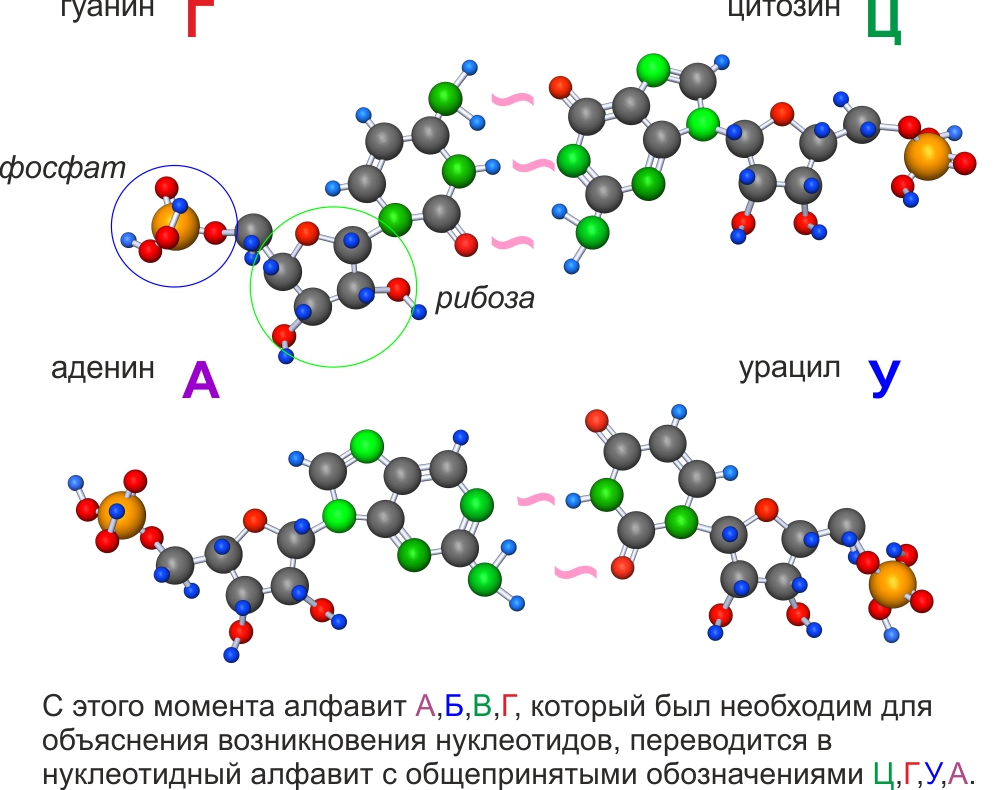
Нуклеотиды, как группа молекул, отличаются от нуклеоноидов только модификацией одной из четырех молекул. Перестав соответствовать Условию №1 для супрамолекул, они утратили возможность самостоятельно супраполимеризоваться, но это компенсировалось созданием более продвинутых рибозимов, способствующих полимеризации.
Молекула, являющаяся полимером 4-х конкретных трехкомпонентых мономеров (нуклеотидов), которые называются аденин, гуанин, урацил и цитозин, независимо от количества мономеров и их последовательности называется рибонуклеиновой кислотой – РНК.

Грязевые котлы – биохимические реакторы
(в этом параграфе и далее обобщение с вариантом формулировки Дарвиновской эволюции и подход к предположению о неферментативной полимеризации)
Выиграв стартовую биохимическую конкуренцию, РНК за счет мутаций начали наследственную эволюцию. Однако для наследственной эволюции требуется много времени, чтобы сменялись многие тысячи и миллионы поколений. Из всех локаций на вулканических островах, на которых могли создаваться нуклеотиды, только грязевые котлы обладали стабильностью достаточной для того, чтобы наследственная эволюция дала результаты.
В грязевых котлах выполнялись следующие условия необходимые для наследственной эволюции, которых нет на остальных локациях являющихся границей трех сред:
-- встроенный источник тепла, сглаживающий влияние неблагоприятных погодных факторов,
-- донор разнообразной химии -- вулкан, извергающий газы и лаву разнообразного химического состава,
-- длительное время существования в стабильном состоянии, время существования грязевых котлов доходит до 10 тысячи лет.
Среднее время существования:

На древней Земле могло быть одновременно около 10 тыс. вулканических островов, и на каждом из них одновременно действовало около 1 тыс. грязевых котлов. Таким образом одновременно на планете действовало около 10 млн. подходящих для наследственной эволюции природных химических реакторов.
|
|
Естественный отбор
Разные модификации РНК, появлявшиеся в грязевых котлах в результате мутаций, начали конкуренцию между собой. Мутации бывают вредными, полезными и нейтральными. Чаще они вредные, т.е. РНК, имеющие такие мутации, проигрывают в конкуренции с РНК не имеющими мутаций и вырождаются. Естественный процесс отсева вредных мутаций и роста в популяции носителей наследственной информации с полезными мутациями называется естественным отбором. Самые удачные последовательности существовали большее количество поколений, создавая свои копии, и подавляли менее удачные варианты.
РНК на пороге жизни
Через некоторое количество поколений происходило разрушение структуры даже самых совершенных РНК в результате негативных внешнего и внутреннего факторов. Внешний -- следствие низкой скорости размножения -- внешняя среда успевала разрушить удачные комбинации быстрее, чем они размножались. Внутренний -- избыточное количество мутаций, которые приводили к вырождению даже успешных вариантов.
Предел Эйгена и скорость размножения
Манфред Эйген доказал, что при большом количестве мутаций количество поколений до распада конечно и вычисляемо. Чтобы количество поколений не имело принципиальных ограничений, количество мутаций должно быть не более одного на поколение -- предел Эйгена. Это биохимический пример реализации принципа, что оптимизация (приспособленность) может происходить одновременно только по одному параметру.
В случае преодоления конкретным видом молекул РНК предела Эйгена и достаточно быстрой скорости размножения, мутации начинают не разрушать РНК, а способствуют приспособлению к изменяющимся внешним условиям за счет случайного возникновения модификаций, некоторые из которых оказываются лучше приспособлены (быстрее размножаются) к изменившимся условиям среды, чем другие. И это есть начало жизни.
Жизнь и дарвиновская эволюция
Жизнь – уровень наследственной эволюции выше порога, который определяется как размножение молекул РНК преодолевших предел Эйгена и размножающихся достаточно быстро, чтобы внешние факторы не успевали разрушать РНК.
Наследственная эволюция, достигшая уровня жизни, называется дарвиновской эволюцией (в честь Чарльза Дарвина, который в середине 19 века открыл и описал в книге "Происхождение видов" существование такой эволюции и ее поразительно разнообразные и поразительно сложные проявления на примере растений и животных ).
Эффективность Дарвиновской эволюции (ДЭ) и сложность ее восприятия
Начинается ли ДЭ (т.е. преодолевают РНК предел Эйгена и достигают достаточно высокой скорости размножения) закономерно или это крайне маловероятно, нам не известно. Перед нами только один пример запуска ДЭ -- зарождение жизни на планете, находящейся в середине пояса жизни желтого карлика по имени Солнце.
Механизм ДЭ прост, но эффективность и результативность ДЭ поразительно высоки. Мутации в последовательности цепочки, несущей наследственную информацию, уничтожение худших вариантов и ускоренное размножение лучших проявляется и закрепляется уже в нескольких поколениях. А когда количество поколений измеряется тысячами и миллионами, достигаются такие выдающиеся результаты в приспособлении к условиям среды, что это кажется невероятным чудом. Тем не менее, это действие только одного механизма -- механизма ДЭ.
Примирить удивительную результативность ДЭ в создании суперсложных биологических механизмов с интуитивным восприятием возможного помогает представление о молекулярных машинах, точнее о молекулярных роботах.
Надо понимать, что сложные одинаковые молекулы не просто являются достаточно точными копиями друг друга, но являются полностью одинаковыми конструкциями с абсолютно одинаковыми физическими и химическими свойствами. Мы не сталкиваемся в реальной жизни с таким не просто очень точным соответствием, а полностью одинаковыми системами, которые при определенных сигналах производят абсолютно одинаковую работу.
Возникают возможности длинных цепочек реакций на сигналы окружающей среды приводящих к абсолютно одинаковым результатам. Обратная связь между изменением (мутацией) и ее результатом абсолютно однозначная, она может быть быстро распознана естественным отбором как полезная, вредная или нейтральная и без задержки и однозначно определит направление эволюции.
Молекулярный робот (МР)
Необходимые для размножения и жизнедеятельности алгоритмы осуществляются разнообразными сложными полимерными молекулами, которые по конструкции и функциональности являются наноаналогами роботов.
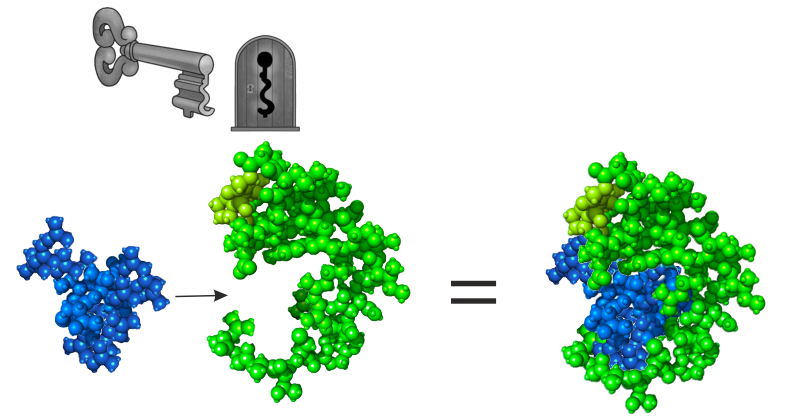
Одно из важнейших свойств МР -- узнавание других молекул не по химическим свойствам, а по пространственным конфигурациям (по принципу ключ - замочная скважина), благодаря чему возникает возможность чрезвычайно специфического влияния на процессы (не достижимое для катализаторов прямого химического действия). Такое свойство является результатом дарвиновского эволюционного совершенствования каждого конкретного механизма структурного катализа.
Кроме структурного катализа МР могли обладать механической функциональностью являющейся реакцией на сигналы среды.
Можно представить МР, который узнает две молекулы, взаимно их ориентирует, а затем их сближает, молекулы реагируют друг с другом, после чего активный центр МР освобождается, и МР многократно проделывает такую работу.
Можно представить МР, который узнает концы обрывков молекулярных цепочек, затем их сближает, концы цепочек связываются, после чего активный центр такого МР освобождается и продолжает работу.
Можно представить МР, который присоединяет к полимерной цепочке новый мономер и делает движение по цепочке за счет энергии высвобождающейся при присоединении этого мономера. Происходит как бы перезарядка МР с одновременным передвижением на один мономер, возникает возможность автоматического срабатывания, приводящего, например, к строительству длинных цепочек из нуклеотидов.
Эволюция, как творец
Механизм ДЭ совершенно лишен творческого начала, это работа алгоритма из двух простых действий -- мутации и естественного отбора работающих на большом количестве поколений последовательностей нуклеотидов, который приводит к совершенствованию МР и созданию новых.
В биологии часто дарвиновскую эволюцию называют просто эволюцией.
Результат эволюции на большом количестве поколений выглядит как целенаправленная творческая работа. И хотя это метафора, часто и оправдано используются словосочетания: Эволюция нашла способ, Эволюция смогла, Эволюция выбрала, Эволюция создала, и вообще Эволюция что-то сделала. Каждый раз надо понимать, что за этим стоит простейший алгоритм, многократное срабатывание которого приводит к описываемому результату.
Аналогичные метафоры используются для продуктов эволюции -- генов, МР и др., например, гены объединились, МР приспособился, вирусы научились и т.п.
|
|
Репликационные полимеразы
Первые МР с явно проявившейся функциональностью были рестриктазы (они разрезали РНК в конкретных местах, разделяя их на отдельные МР -- протогены), лигазы (они сшивали концы обрывков цепочек) и репликационные полимеразы (они ускоряли строительство цепочки из нуклеотидов). Первые МР имели длину порядка от 100 для лигазы до 200 нуклеотидов для полимеразы и состояли только из последовательности нуклеотидов (без интеграции других молекул в состав МР).
За какое-то время до Второй тяжелой бомбардировки РНК с помощью создания МР (самым сложным и необходимым из которых была полимераза) достигли скорости и точности размножения достаточно высокого уровня сложности для запуска ДЭ, т.е. для запуска жизни. И уже благодаря ДЭ жизнь, создав множество МР и все время повышая их сложность и совершенство, достигла такого уровня, что смогла пережить Вторую тяжелую бомбардировку.
Единичный грязевой котел -- колыбель жизни
Если образование РНК могло произойти параллельно во многих лужах и грязевых котлах, то вся эволюция, начиная от первых рибозимов и до начала ДЭ, могла произойти только в одном грязевом котле, или, при более оптимистическом взгляде, в нескольких. Это следствие крайне малой вероятности появления полимеразы для достаточно высокой скорости размножения.
Проблема дополимеразной полимеризации нуклеотидов
Процесс образования нуклеотидов и РНК в поясе жизни химически закономерен и на разных планетах на расстоянии от 100 до 250 млн.км от своих желтых карликов, скорее всего, происходит одинаково. Учеными не найдены отличные от нуклеотидов, хотя бы теоретически правдоподобные варианты биомономеров обладающих свойствами необходимыми для создания полимера жизни. Если жизнь еще где-нибудь во Вселенной существует, то в своем основании она имеет нуклеотиды полимеризующиеся в РНК.
Преодолевается ли предел Эйгена (одна мутация на одну репликацию) при достаточной скорости репликации, и начинается ДЭ закономерно, или это крайне маловероятно, нам не известно, мы знаем только один пример зарождения жизни.
Возможность преодоления предела Эйгена фрагментами РНК при определенном уровне и частоте УФ и температурных воздействий (необходимо довести количество мутаций при репликации от 1 на 50 до 1 на 200) лабораторно установлена. Большей сложностью является низкая скорость размножения.
Лабораторно, с применением самых разнообразных условий, из нуклеотидов не удается получать сколько-нибудь длинных цепочек. Достигаются цепочки максимум в 20 нуклеотидов, после чего цепочки рассыпаются.
Наука считает, что для выхода на Дарвиновскую эволюцию необходимо, чтобы функциональность одного фрагмента цепи РНК достигла уровня полимеразы, т.е. такого сложного и специфического молекулярного робота, который сам сможет сшивать отдельные нуклеотиды (причем желательно с уменьшением количества мутаций по сравнению с самопроизвольным сшиванием), многократно ускоряя размножение.
С появлением полимераз размножение смогло стать намного более частым, чем раз в сутки, т.е. оторвалось от циклического суточного УФ и температурного воздействия, как необходимых факторов механизма размножения.
Полимераза является необходимым этапом, но объяснение достижения РНК такого уровня сложности, как полимераза, пока науке не удается. Чрезвычайно низковероятное соединение в правильной последовательности необходимых для полимеразы более 200 нуклеотидов в условиях низкой скорости полимеризации делает даже разовое такое событие крайне маловероятном для всей Вселенной за все время ее существования.
Для объяснения возникновения полимеразы учеными привлекается теория бесконечного количества вселенных, в одной из которых тогда и может возникнуть жизнь (бесконечно малая вероятность умножается на бесконечное количество попыток). Другие ученые считают, что достижение РНК сложности уровня полимеразы закономерно, они ищут пути ее возникновения через дополимеразные механизмы запуска Дарвиновской эволюции. Подробнее здесь.
Нуклеоноиды как дополимеразные молекулы жизни
Объяснение дополимеразной эволюции РНК может базироваться на гипотезе о предшествующих нуклеотидам нуклеоноидам, которые, создавая по три поляризационные связи в каждой паре и более успешно объединяясь в супрамолекулы, могли полимеризоваться с большей скоростью и в более длинные цепочки без помощи полимеразы. В этом случае началом жизни нужно считать появление нуклеоноидов способных к самостоятельной дарвиновской эволюции. Для подтверждения гипотезы нуклеоноидов необходимы эксперименты по неферментативной (без участия полимеразы) полимеризации нуклеоноидов.
|
|
|